

Structural basis of Nrd1-Nab3 heterodimerization
- PMID: 35022249
- PMCID: PMC8761494
- DOI: 10.26508/lsa.202101252
Structural basis of Nrd1-Nab3 heterodimerization
Abstract
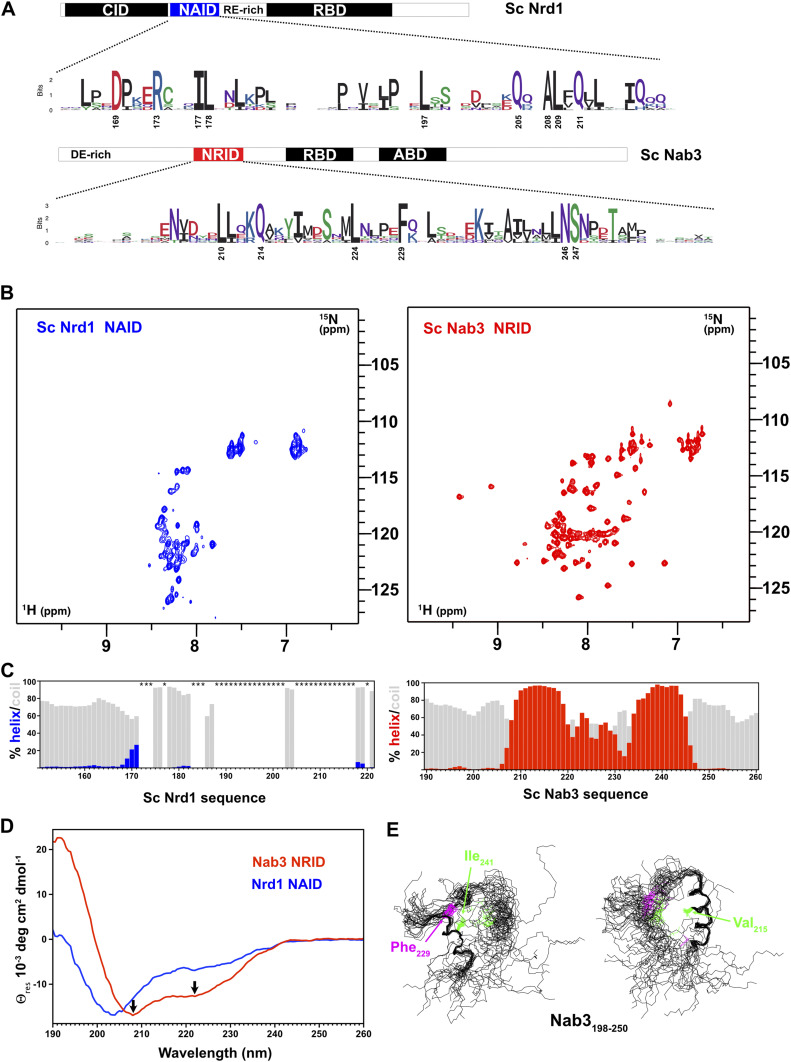
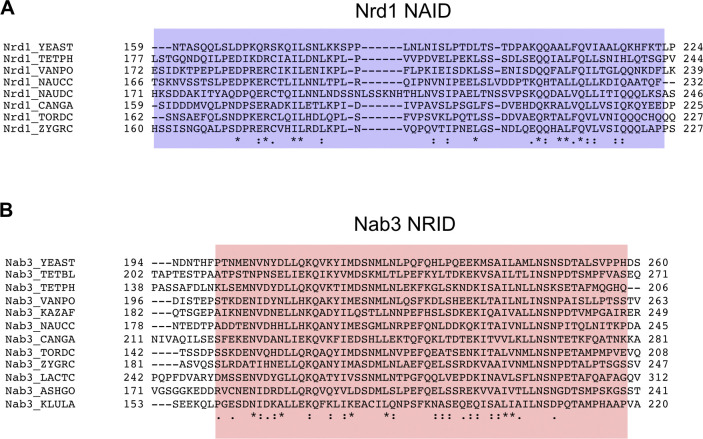
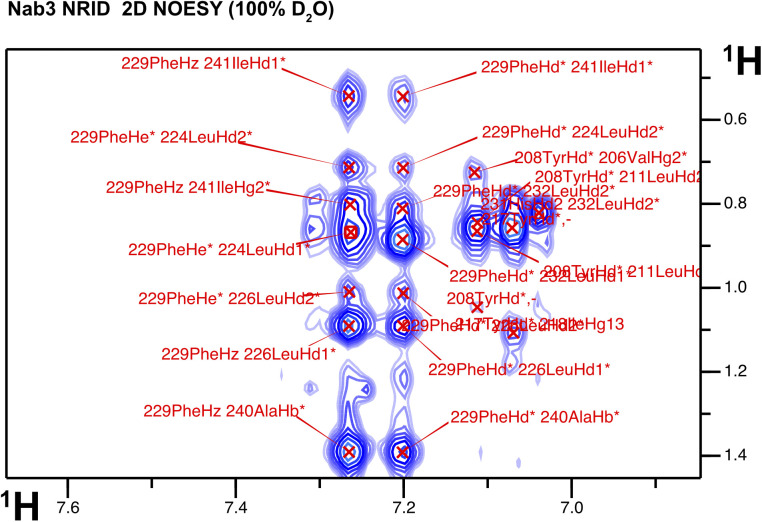
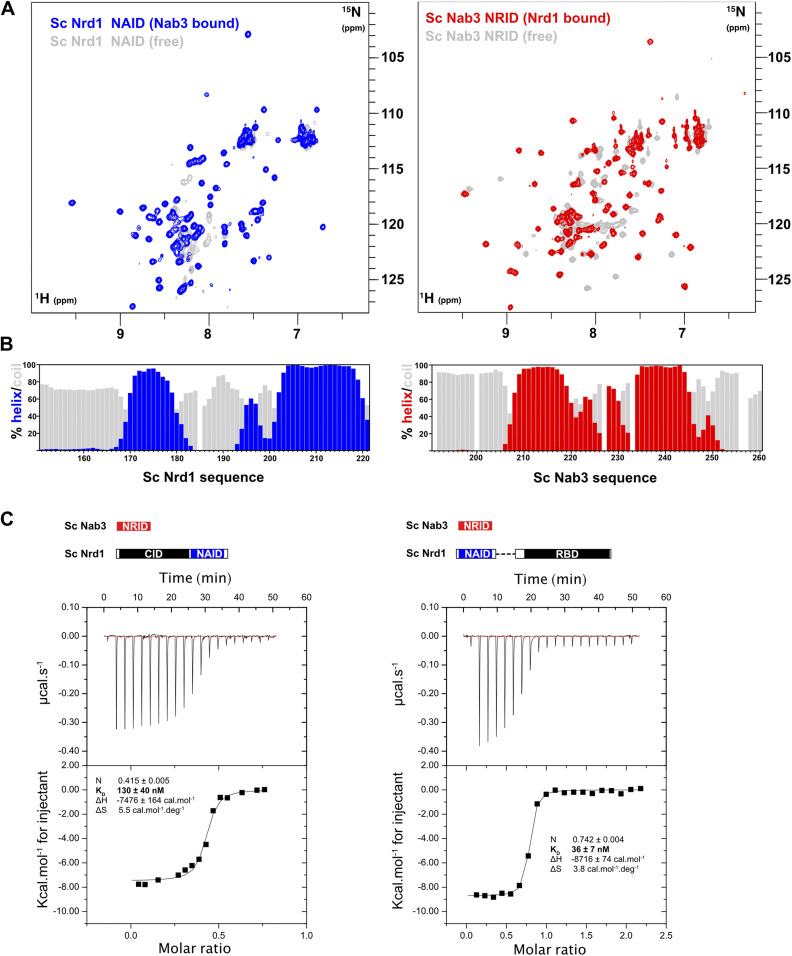
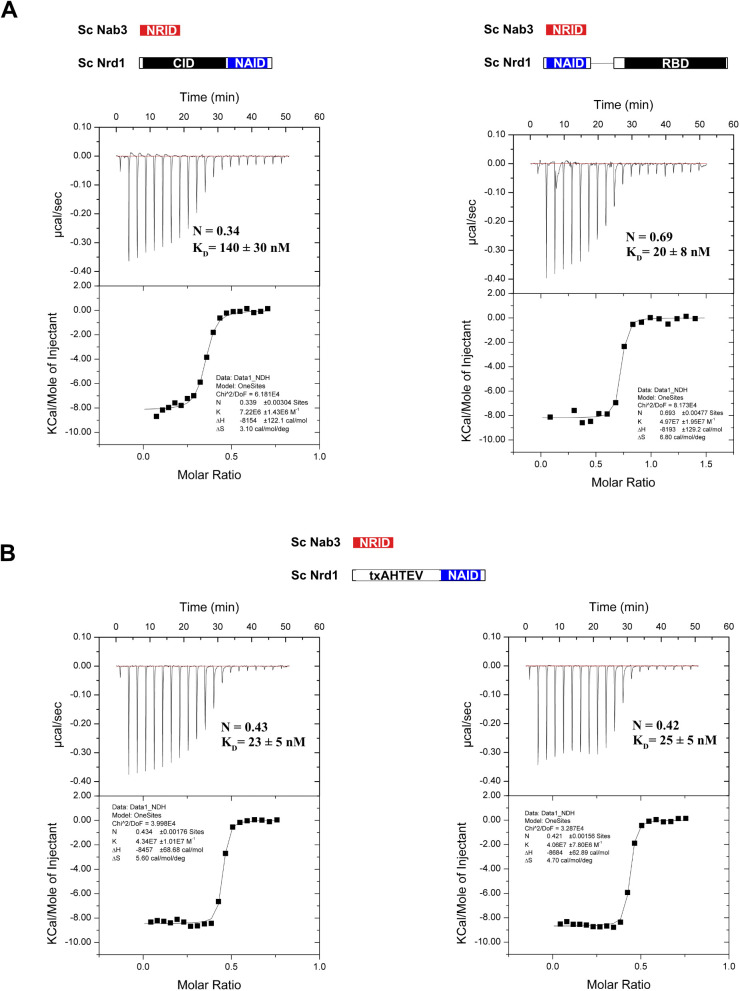
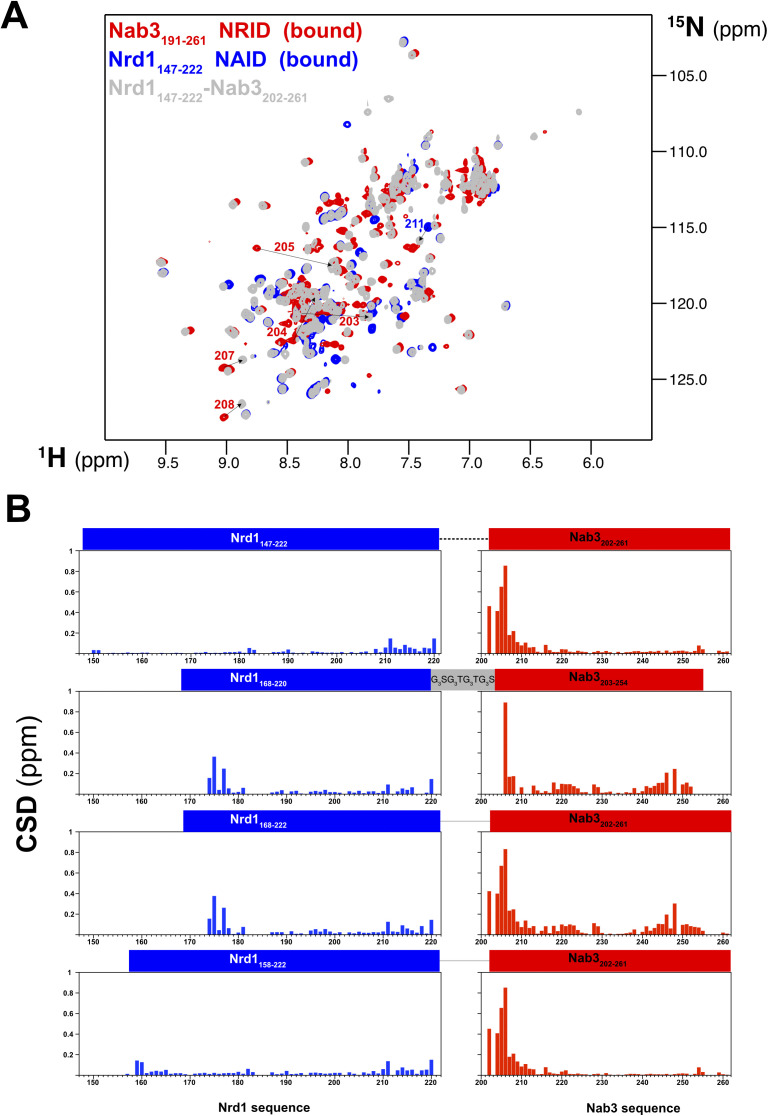
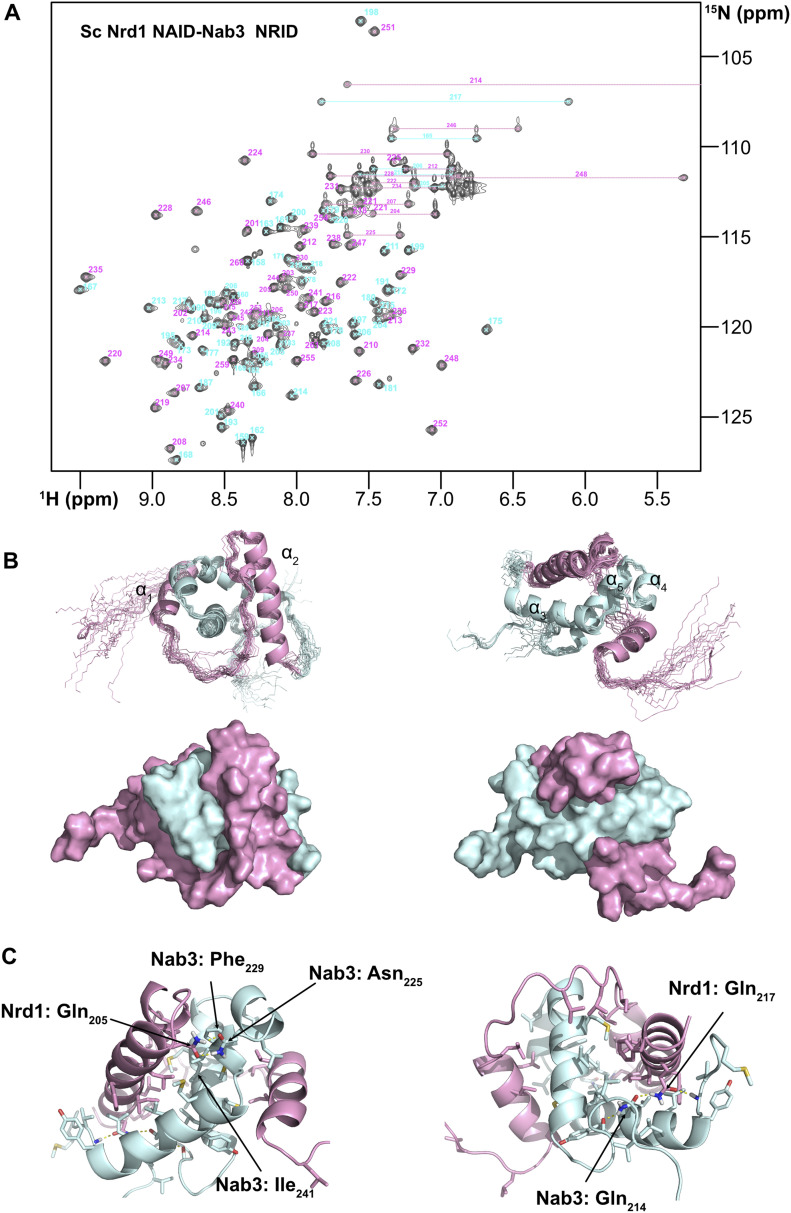
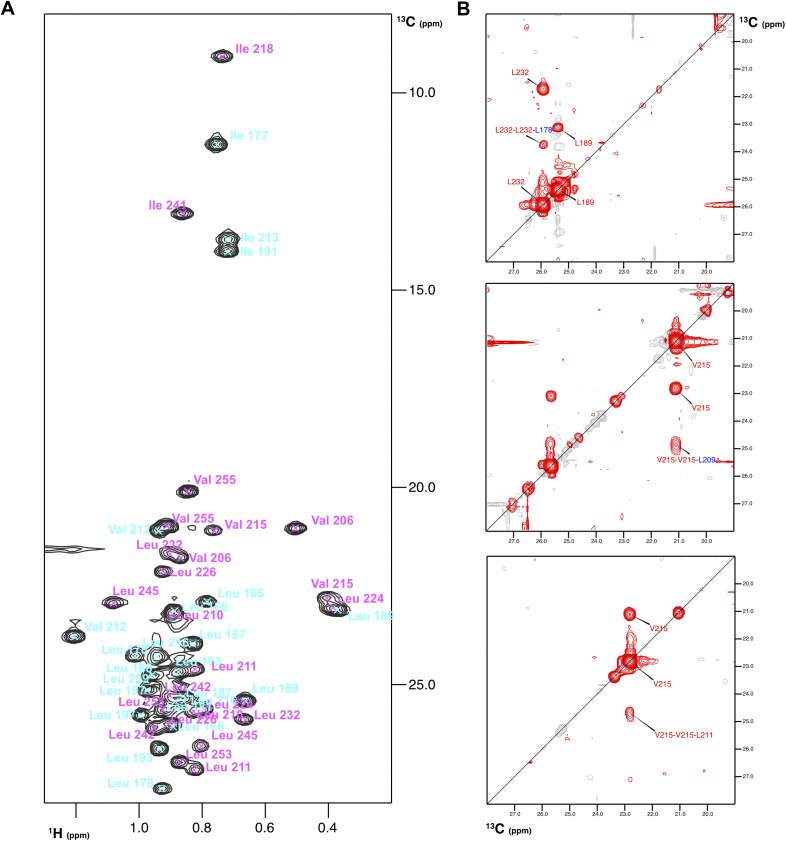
Heterodimerization of RNA binding proteins Nrd1 and Nab3 is essential to communicate the RNA recognition in the nascent transcript with the Nrd1 recognition of the Ser5-phosphorylated Rbp1 C-terminal domain in RNA polymerase II. The structure of a Nrd1-Nab3 chimera reveals the basis of heterodimerization, filling a missing gap in knowledge of this system. The free form of the Nrd1 interaction domain of Nab3 (NRID) forms a multi-state three-helix bundle that is clamped in a single conformation upon complex formation with the Nab3 interaction domain of Nrd1 (NAID). The latter domain forms two long helices that wrap around NRID, resulting in an extensive protein-protein interface that would explain the highly favorable free energy of heterodimerization. Mutagenesis of some conserved hydrophobic residues involved in the heterodimerization leads to temperature-sensitive phenotypes, revealing the importance of this interaction in yeast cell fitness. The Nrd1-Nab3 structure resembles the previously reported Rna14/Rna15 heterodimer structure, which is part of the poly(A)-dependent termination pathway, suggesting that both machineries use similar structural solutions despite they share little sequence homology and are potentially evolutionary divergent.
© 2022 Chaves-Arquero et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
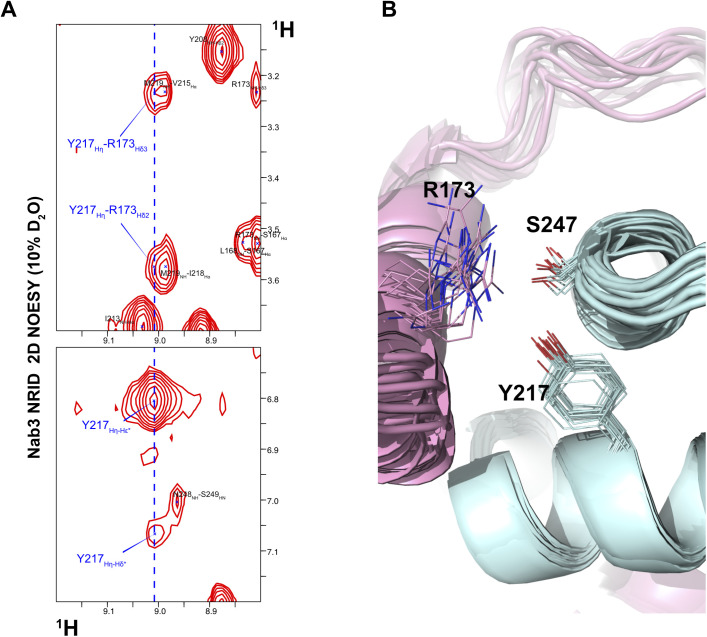
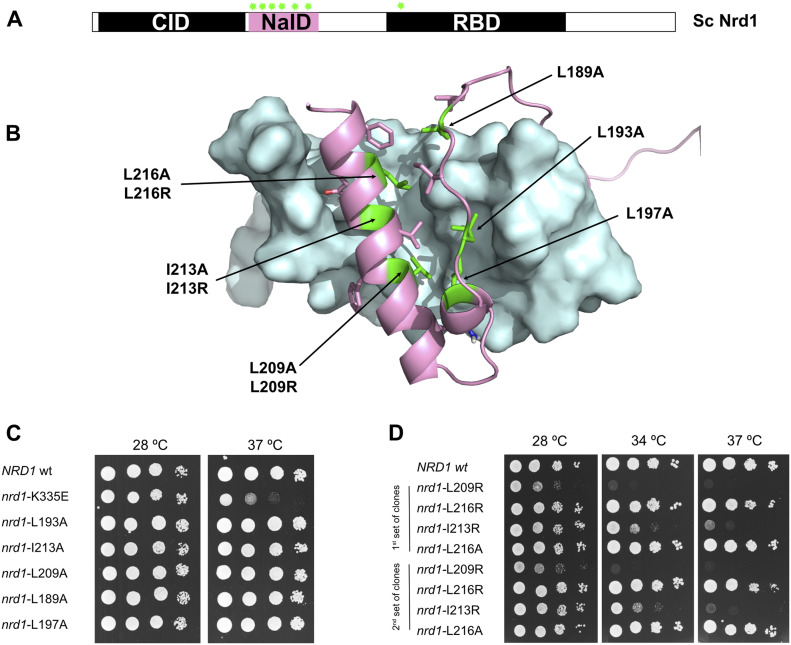
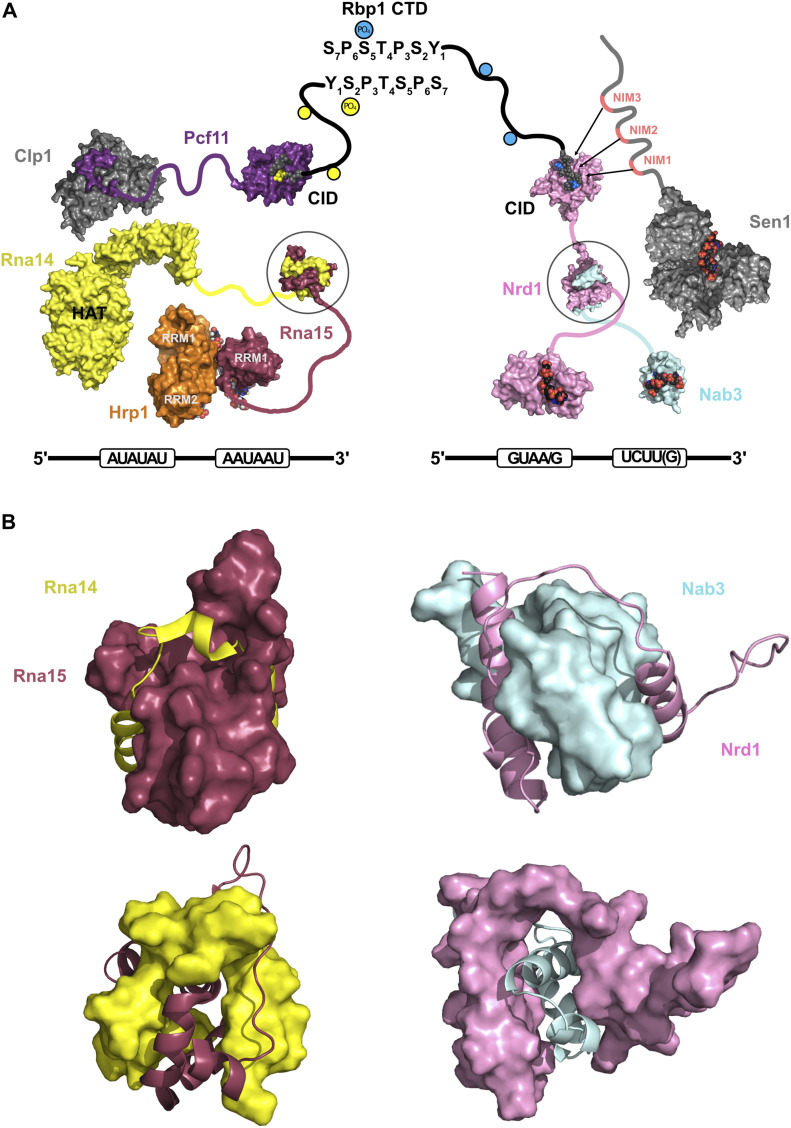
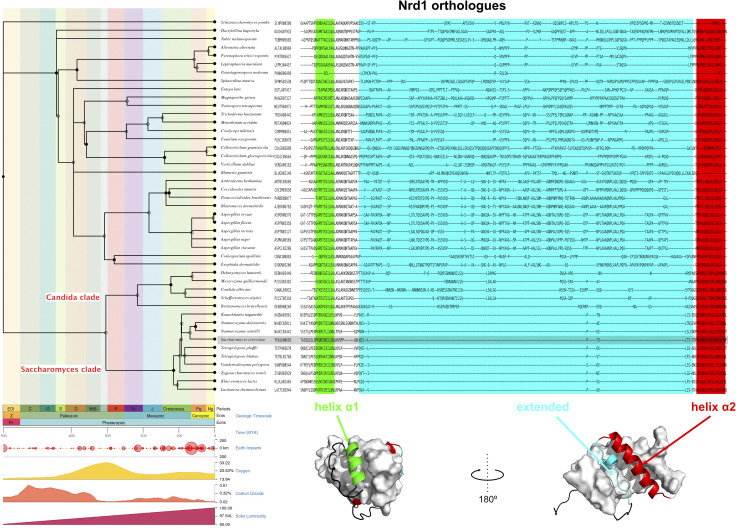
References
-
- Altenhoff AM, Train CM, Gilbert KJ, Mediratta I, Mendes De Farias T, Moi D, Nevers Y, Radoykova HS, Rossier V, Warwick Vesztrocy A, et al. (2020) OMA orthology in 2021: Website overhaul, conserved isoforms, ancestral gene order and more. Nucleic Acids Res 49: 373–379. 10.1093/nar/gkaa1007 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous