

SARS-CoV-2 mRNA vaccination elicits a robust and persistent T follicular helper cell response in humans
- PMID: 35026152
- PMCID: PMC8695127
- DOI: 10.1016/j.cell.2021.12.026
SARS-CoV-2 mRNA vaccination elicits a robust and persistent T follicular helper cell response in humans
Abstract
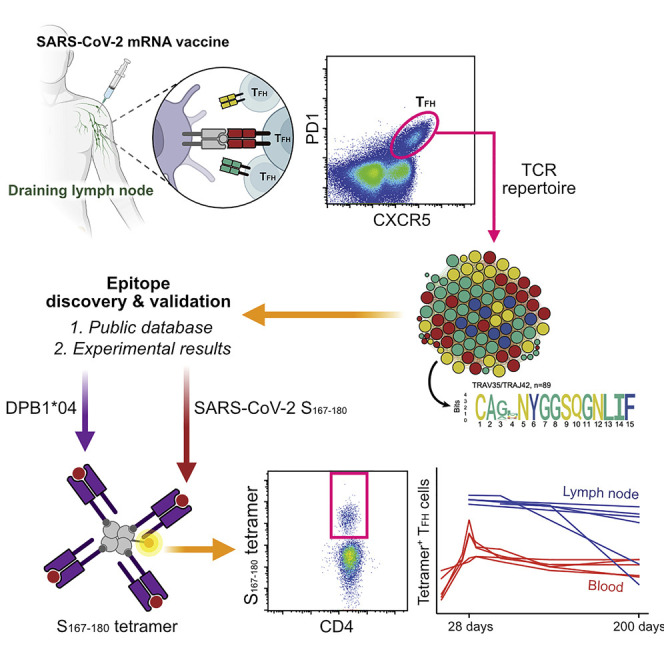
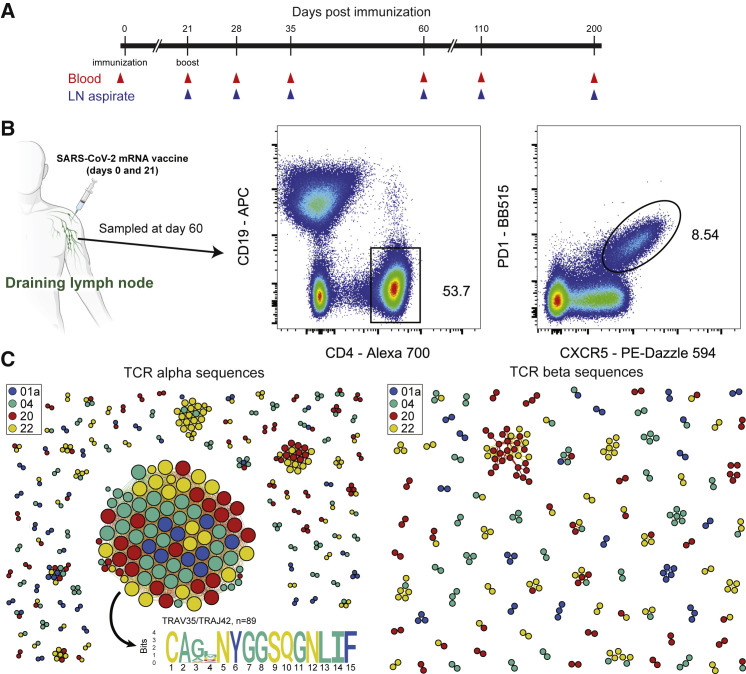
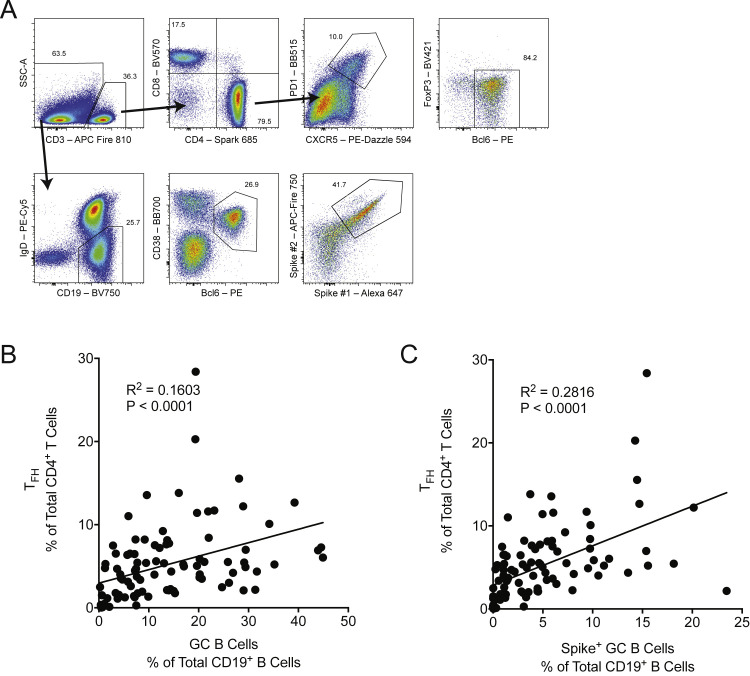
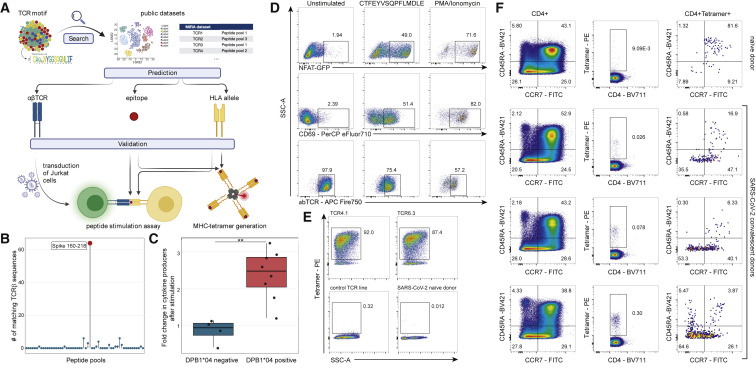
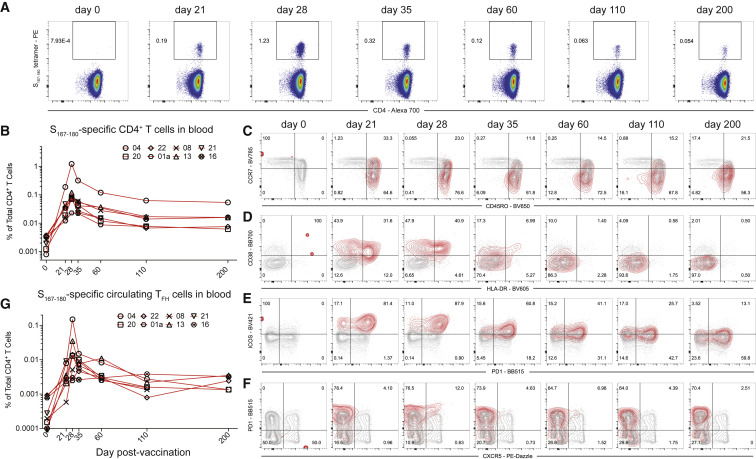
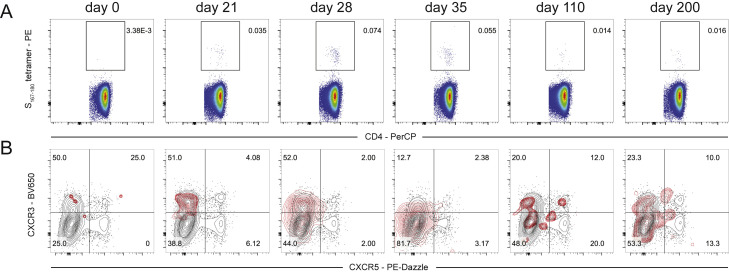
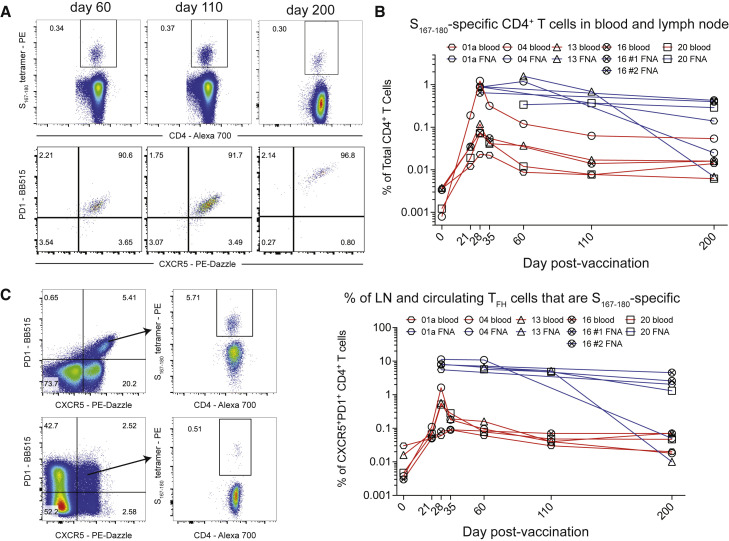
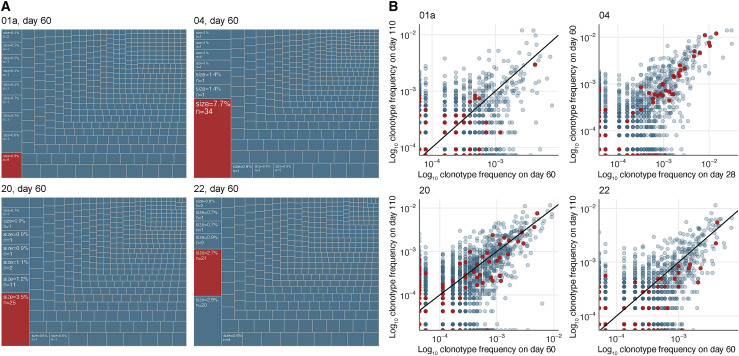
SARS-CoV-2 mRNA vaccines induce robust anti-spike (S) antibody and CD4+ T cell responses. It is not yet clear whether vaccine-induced follicular helper CD4+ T (TFH) cell responses contribute to this outstanding immunogenicity. Using fine-needle aspiration of draining axillary lymph nodes from individuals who received the BNT162b2 mRNA vaccine, we evaluated the T cell receptor sequences and phenotype of lymph node TFH. Mining of the responding TFH T cell receptor repertoire revealed a strikingly immunodominant HLA-DPB1∗04-restricted response to S167-180 in individuals with this allele, which is among the most common HLA alleles in humans. Paired blood and lymph node specimens show that while circulating S-specific TFH cells peak one week after the second immunization, S-specific TFH persist at nearly constant frequencies for at least six months. Collectively, our results underscore the key role that robust TFH cell responses play in establishing long-term immunity by this efficacious human vaccine.
Keywords: CD4(+) T cell; COVID-19; SARS-CoV-2; T follicular helper cell; TCR repertoire; human immunology; lymph node; mRNA vaccination.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The Ellebedy laboratory received funding under sponsored research agreements that are unrelated to the data presented in the current study from Emergent BioSolutions and from AbbVie. A.H.E. has received consulting payments from Mubadala Investment Company, InBios International, and Fimbrion Therapeutics and is the founder of ImmuneBio Consulting. P.G.T. has consulted and/or received honoraria and travel support from Illumina, Johnson and Johnson, and 10X Genomics. P.G.T. serves on the Scientific Advisory Board of Immunoscape and Cytoagents. The authors have applied for patents covering some aspects of these studies.
Figures
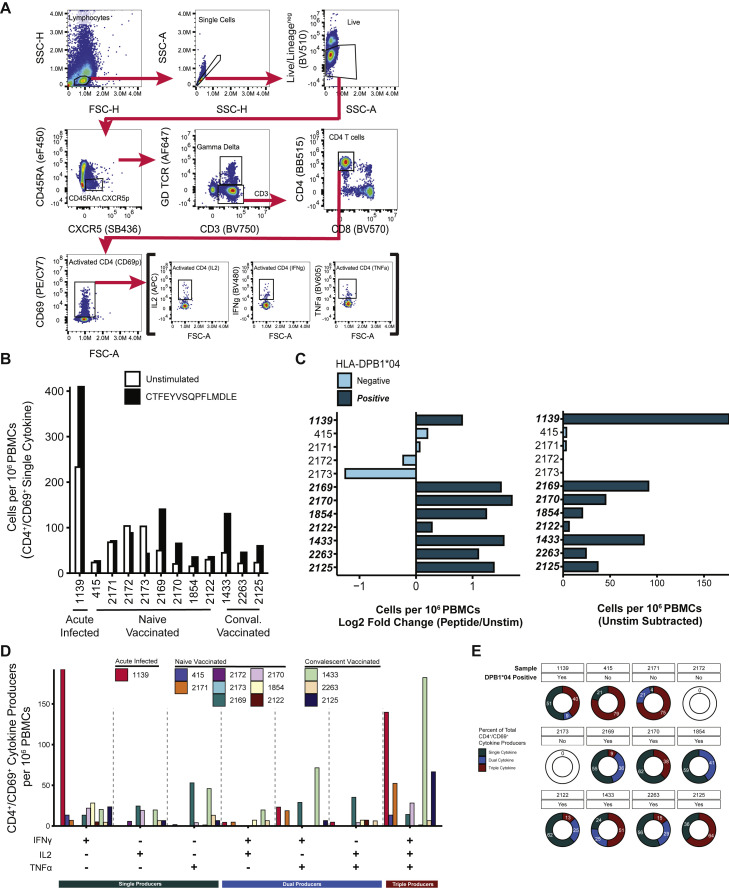
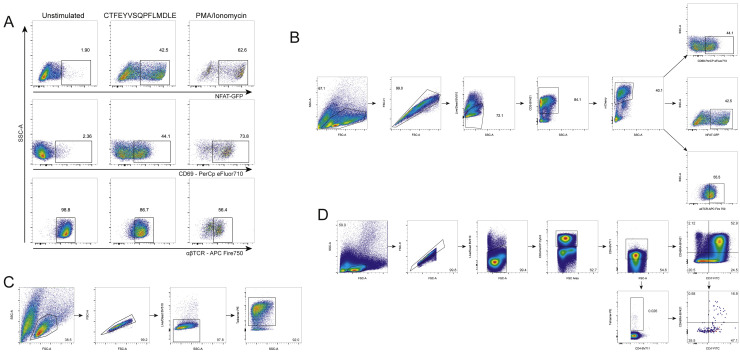
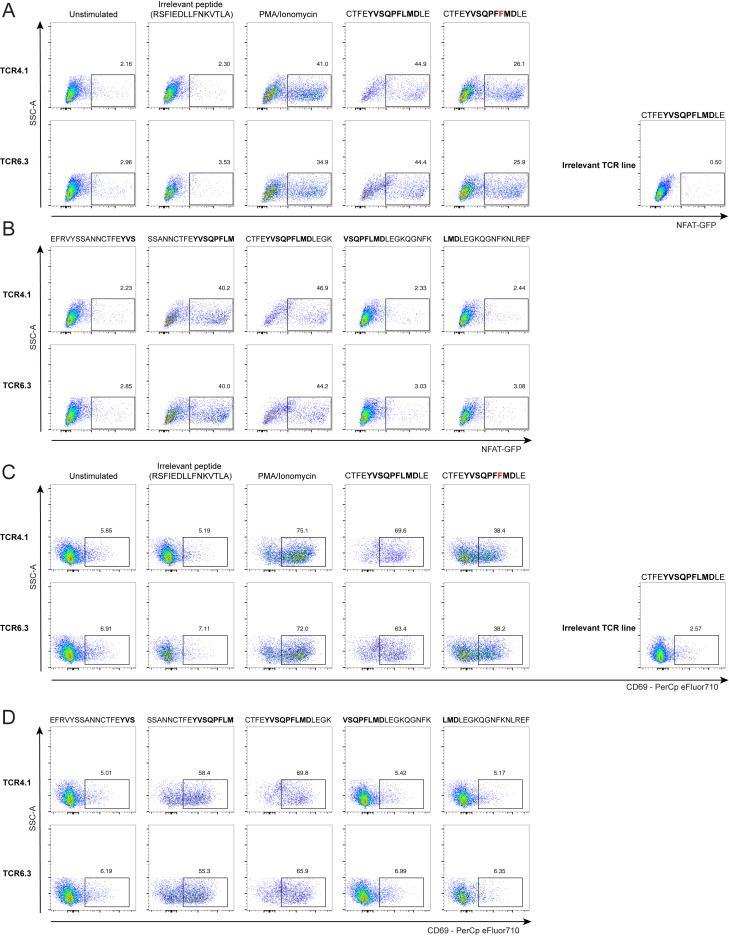
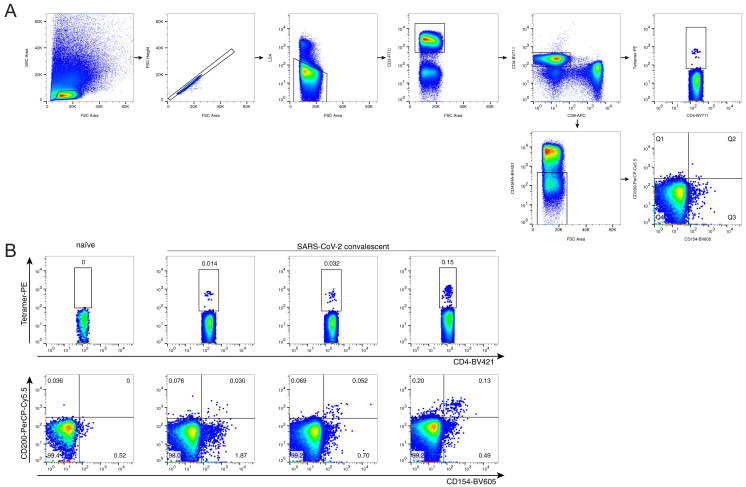
References
-
- Bolotin D.A., Poslavsky S., Mitrophanov I., Shugay M., Mamedov I.Z., Putintseva E.V., Chudakov D.M. MiXCR: software for comprehensive adaptive immunity profiling. Nat. Methods. 2015;12:380–381. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
