

Rewards of divergence in sequences, 3-D structures and dynamics of yeast and human spliceosome SF3b complexes
- PMID: 35028595
- PMCID: PMC8714771
- DOI: 10.1016/j.crstbi.2021.05.003
Rewards of divergence in sequences, 3-D structures and dynamics of yeast and human spliceosome SF3b complexes
Abstract
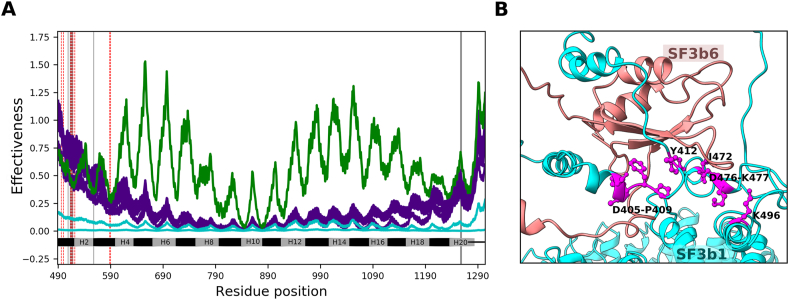
The evolution of homologous and functionally equivalent multiprotein assemblies is intriguing considering sequence divergence of constituent proteins. Here, we studied the implications of protein sequence divergence on the structure, dynamics and function of homologous yeast and human SF3b spliceosomal subcomplexes. Human and yeast SF3b comprise of 7 and 6 proteins respectively, with all yeast proteins homologous to their human counterparts at moderate sequence identity. SF3b6, an additional component in the human SF3b, interacts with the N-terminal extension of SF3b1 while the yeast homologue Hsh155 lacks the equivalent region. Through detailed homology studies, we show that SF3b6 is absent not only in yeast but in multiple lineages of eukaryotes implying that it is critical in specific organisms. We probed for the potential role of SF3b6 in the spliceosome assembled form through structural and flexibility analyses. By analysing normal modes derived from anisotropic network models of SF3b1, we demonstrate that when SF3b1 is bound to SF3b6, similarities in the magnitude of residue motions (0.86) and inter-residue correlated motions (0.94) with Hsh155 are significantly higher than when SF3b1 is considered in isolation (0.21 and 0.89 respectively). We observed that SF3b6 promotes functionally relevant 'open-to-close' transition in SF3b1 by enhancing concerted residue motions. Such motions are found to occur in the Hsh155 without SF3b6. The presence of SF3b6 influences motions of 16 residues that interact with U2 snRNA/branchpoint duplex and supports the participation of its interface residues in long-range communication in the SF3b1. These results advocate that SF3b6 potentially acts as an allosteric regulator of SF3b1 for BPS selection and might play a role in alternative splicing. Furthermore, we observe variability in the relative orientation of SF3b4 and in the local structure of three β-propeller domains of SF3b3 with reference to their yeast counterparts. Such differences influence the inter-protein interactions of SF3b between these two organisms. Together, our findings highlight features of SF3b evolution and suggests that the human SF3b may have evolved sophisticated mechanisms to fine tune its molecular function.
Keywords: Allostery; BPS, branch-point sequence; Bact, activated B spliceosome assembly; Cryo-EM structure; Cryo-EM, cryo-electron microscopy; DOPE, discrete optimized protein energy; NMA, normal mode analysis; PDB, protein data bank; Protein dynamics; RMSD, root mean square deviation; RRM, RNA recognition motif; SF3b complex; SF3b1; SF3b1SF3b6−bound, SF3b1 bound to SF3b6; SF3b1iso, SF3b1 in isolation; SIP, square inner product; Spliceosome.
© 2021 The Authors.
Conflict of interest statement
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures







References
-
- Bai R., Wan R., Yan C., Lei J., Shi Y. Structures of the fully assembled Saccharomyces cerevisiae spliceosome before activation. Science. 2018;360:1423–1429. - PubMed
-
- Berman H.M., Battistuz T., Bhat T.N., Bluhm W.F., Bourne P.E., Burkhardt K., Feng Z., Gilliland G.L., Iype L., Jain S., et al. The protein data bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002;58:899–907. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
