

A nuclear function for an oncogenic microRNA as a modulator of snRNA and splicing
- PMID: 35033060
- PMCID: PMC8760648
- DOI: 10.1186/s12943-022-01494-z
A nuclear function for an oncogenic microRNA as a modulator of snRNA and splicing
Abstract
Background: miRNAs are regulatory transcripts established as repressors of mRNA stability and translation that have been functionally implicated in carcinogenesis. miR-10b is one of the key onco-miRs associated with multiple forms of cancer. Malignant gliomas exhibit particularly striking dependence on miR-10b. However, despite the therapeutic potential of miR-10b targeting, this miRNA's poorly investigated and largely unconventional properties hamper the clinical translation.
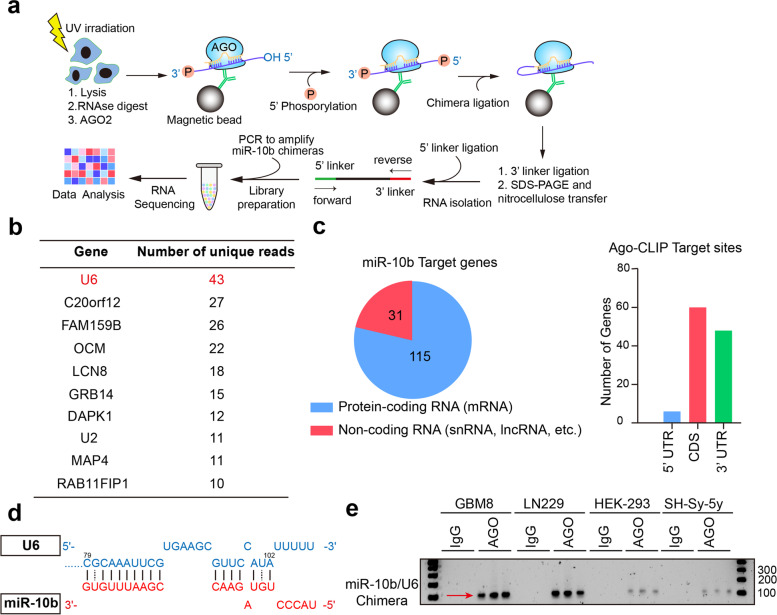
Methods: We utilized Covalent Ligation of Endogenous Argonaute-bound RNAs and their high-throughput RNA sequencing to identify miR-10b interactome and a combination of biochemical and imaging approaches for target validation. They included Crosslinking and RNA immunoprecipitation with spliceosomal proteins, a combination of miRNA FISH with protein immunofluorescence in glioma cells and patient-derived tumors, native Northern blotting, and the transcriptome-wide analysis of alternative splicing.
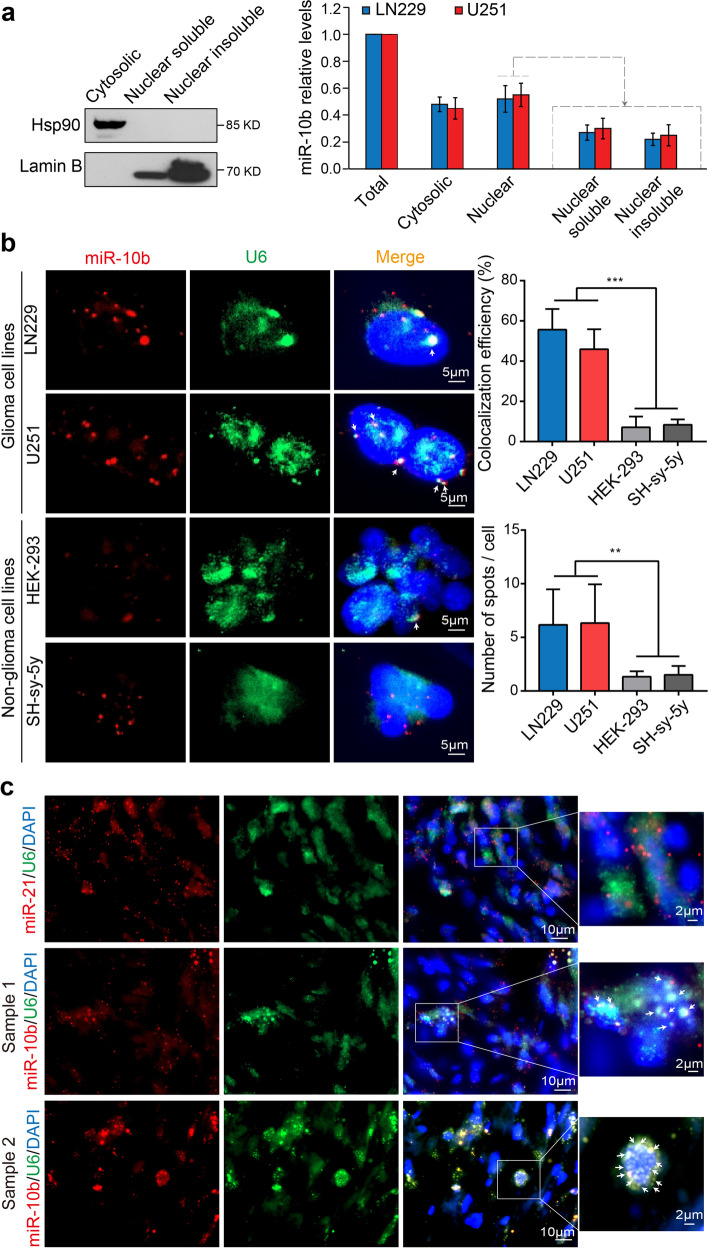
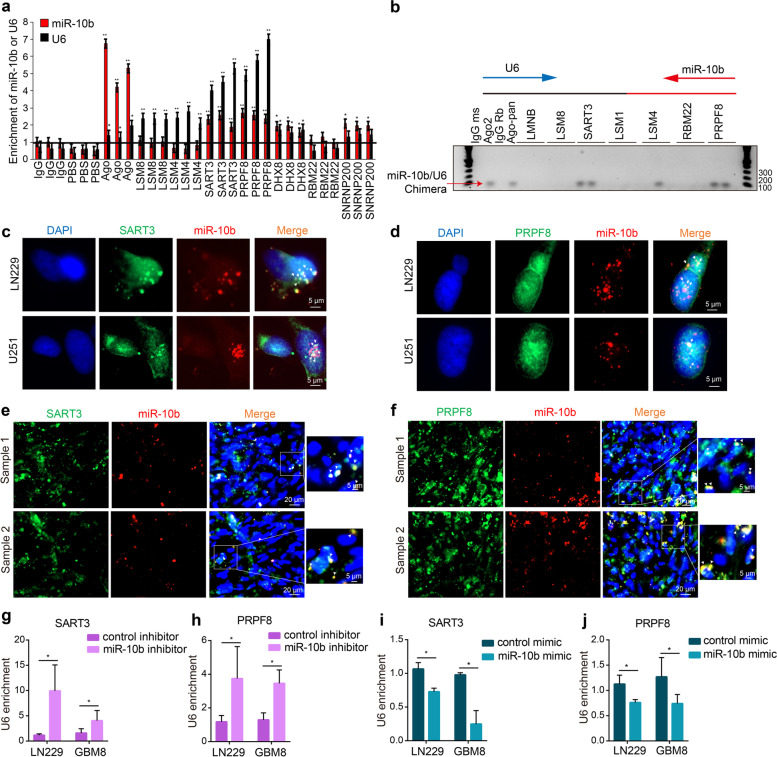
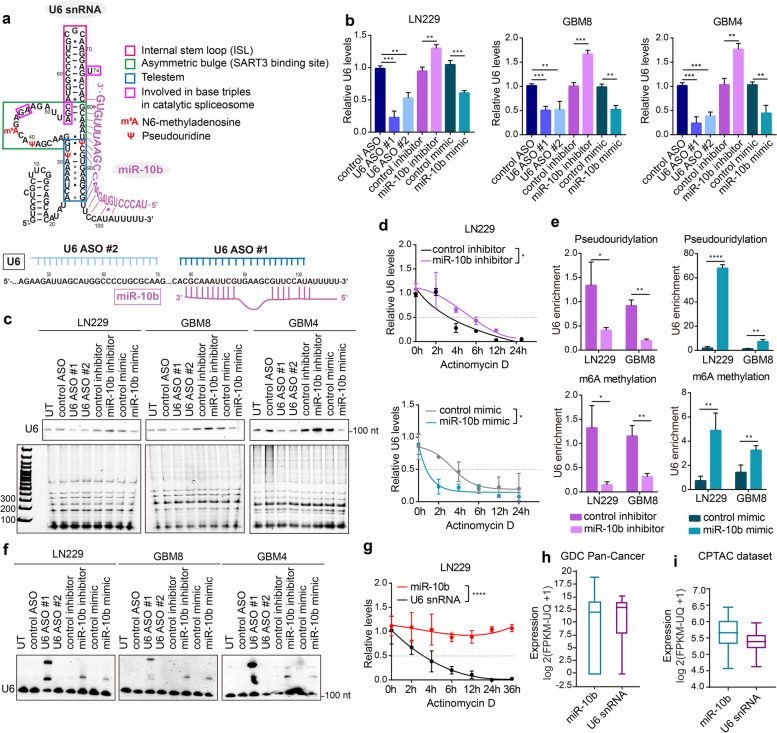
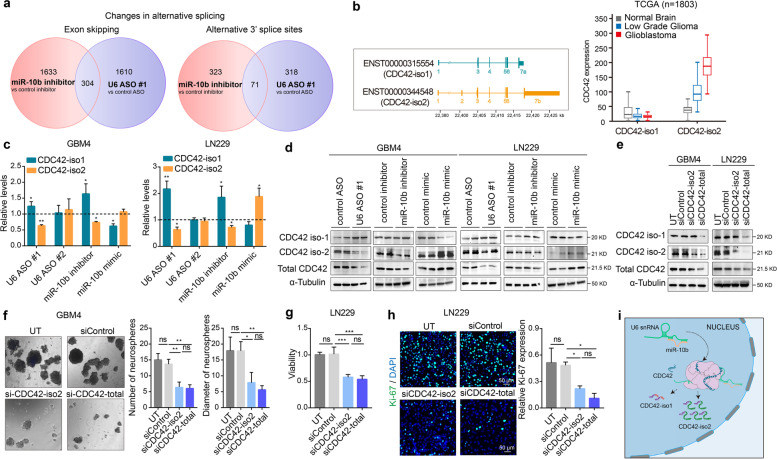
Results: We demonstrate that miR-10b binds to U6 snRNA, a core component of the spliceosomal machinery. We provide evidence of the direct binding between miR-10b and U6, in situ imaging of miR-10b and U6 co-localization in glioma cells and tumors, and biochemical co-isolation of miR-10b with the components of the spliceosome. We further demonstrate that miR-10b modulates U6 N-6-adenosine methylation and pseudouridylation, U6 binding to splicing factors SART3 and PRPF8, and regulates U6 stability, conformation, and levels. These effects on U6 result in global splicing alterations, exemplified by the altered ratio of the isoforms of a small GTPase CDC42, reduced overall CDC42 levels, and downstream CDC42 -mediated effects on cell viability.
Conclusions: We identified U6 snRNA, the key RNA component of the spliceosome, as the top miR-10b target in glioblastoma. We, therefore, present an unexpected intersection of the miRNA and splicing machineries and a new nuclear function for a major cancer-associated miRNA.
Keywords: CDC42; Glioblastoma; Nucleus; Splicing machinery; U6 snRNA; miR-10b.
© 2022. The Author(s).
Conflict of interest statement
All other authors declare no competing financial interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
