

Characterization of the SARS-CoV-2 ExoN (nsp14ExoN-nsp10) complex: implications for its role in viral genome stability and inhibitor identification
- PMID: 35037045
- PMCID: PMC8860572
- DOI: 10.1093/nar/gkab1303
Characterization of the SARS-CoV-2 ExoN (nsp14ExoN-nsp10) complex: implications for its role in viral genome stability and inhibitor identification
Abstract
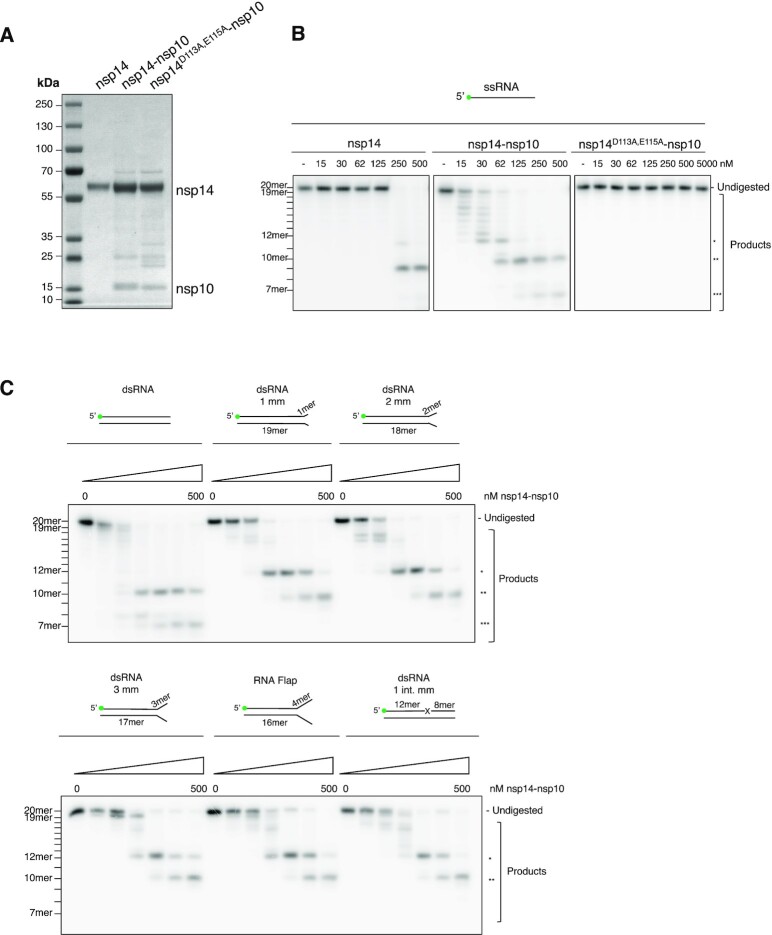
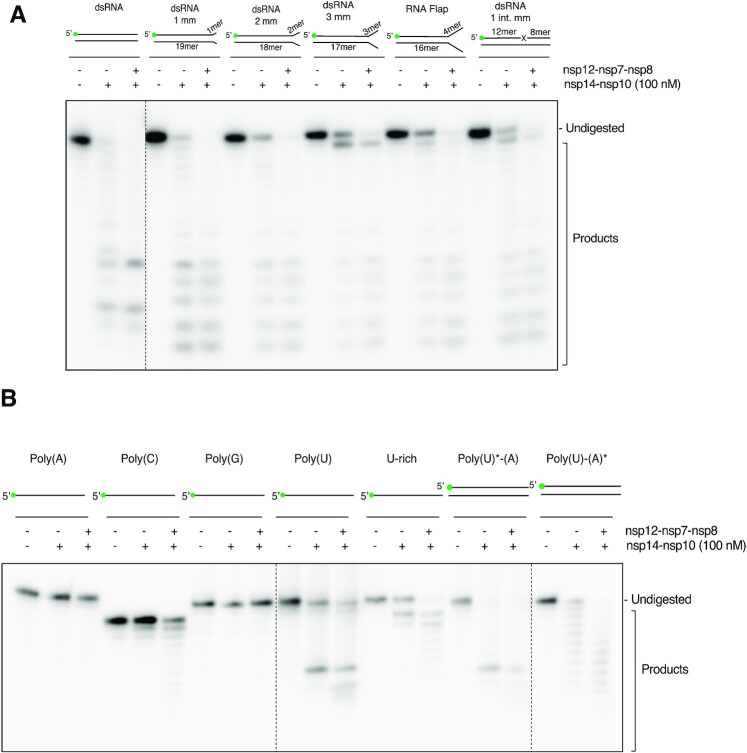
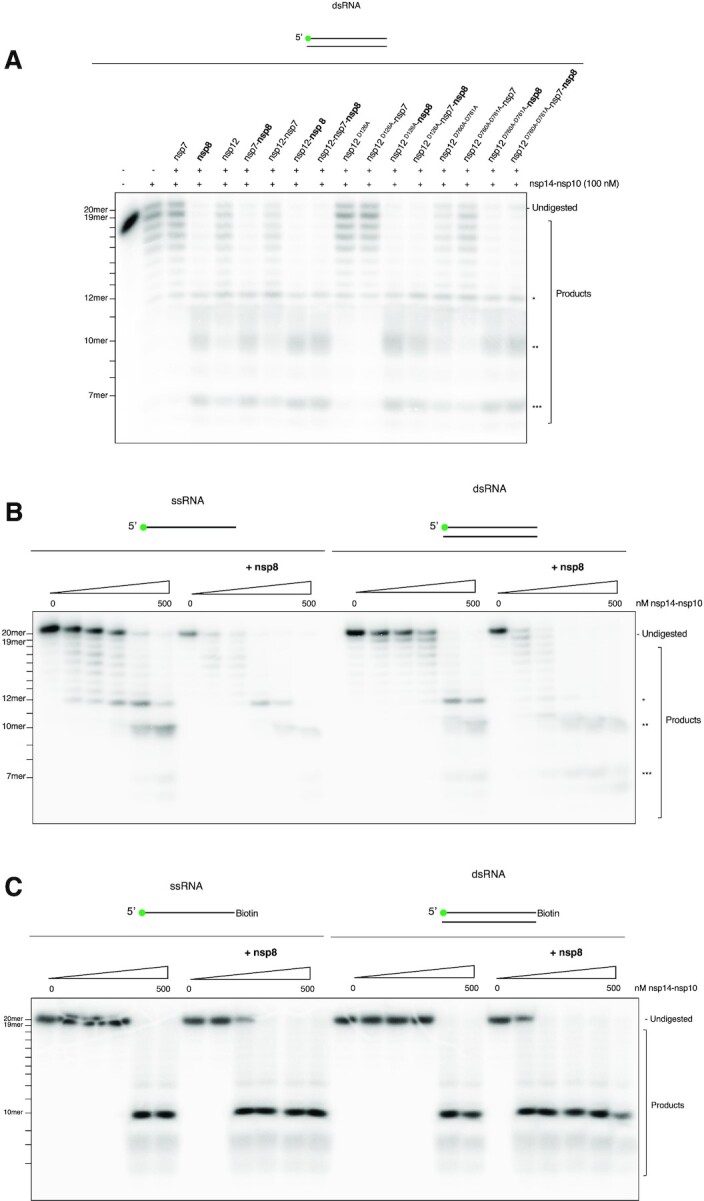
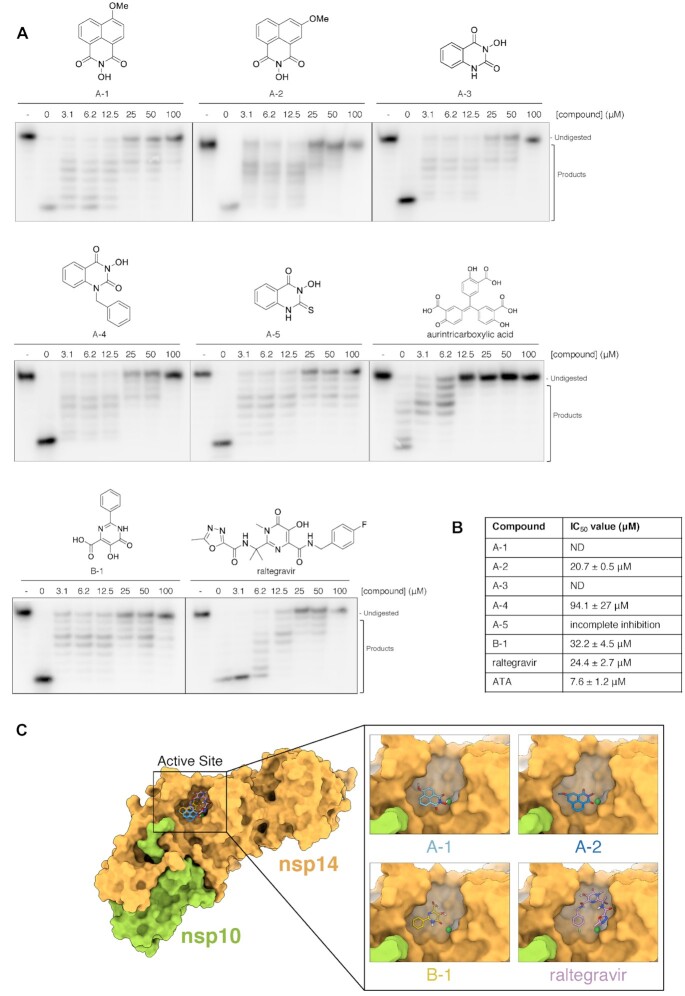
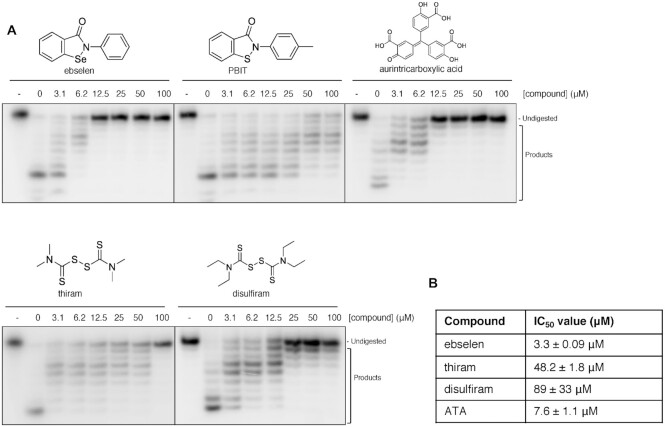
The SARS-CoV-2 coronavirus is the causal agent of the current global pandemic. SARS-CoV-2 belongs to an order, Nidovirales, with very large RNA genomes. It is proposed that the fidelity of coronavirus (CoV) genome replication is aided by an RNA nuclease complex, comprising the non-structural proteins 14 and 10 (nsp14-nsp10), an attractive target for antiviral inhibition. Our results validate reports that the SARS-CoV-2 nsp14-nsp10 complex has RNase activity. Detailed functional characterization reveals nsp14-nsp10 is a versatile nuclease capable of digesting a wide variety of RNA structures, including those with a blocked 3'-terminus. Consistent with a role in maintaining viral genome integrity during replication, we find that nsp14-nsp10 activity is enhanced by the viral RNA-dependent RNA polymerase complex (RdRp) consisting of nsp12-nsp7-nsp8 (nsp12-7-8) and demonstrate that this stimulation is mediated by nsp8. We propose that the role of nsp14-nsp10 in maintaining replication fidelity goes beyond classical proofreading by purging the nascent replicating RNA strand of a range of potentially replication-terminating aberrations. Using our developed assays, we identify drug and drug-like molecules that inhibit nsp14-nsp10, including the known SARS-CoV-2 major protease (Mpro) inhibitor ebselen and the HIV integrase inhibitor raltegravir, revealing the potential for multifunctional inhibitors in COVID-19 treatment.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures

References
-
- Fung T.S., Liu D.X.. Human coronavirus: host-pathogen interaction. Annu Rev Microbiol. 2019; 73:529–557. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
