

CRISPR Interference Reveals That All- Trans-Retinoic Acid Promotes Macrophage Control of Mycobacterium tuberculosis by Limiting Bacterial Access to Cholesterol and Propionyl Coenzyme A
- PMID: 35038923
- PMCID: PMC8764544
- DOI: 10.1128/mbio.03683-21
CRISPR Interference Reveals That All- Trans-Retinoic Acid Promotes Macrophage Control of Mycobacterium tuberculosis by Limiting Bacterial Access to Cholesterol and Propionyl Coenzyme A
Abstract
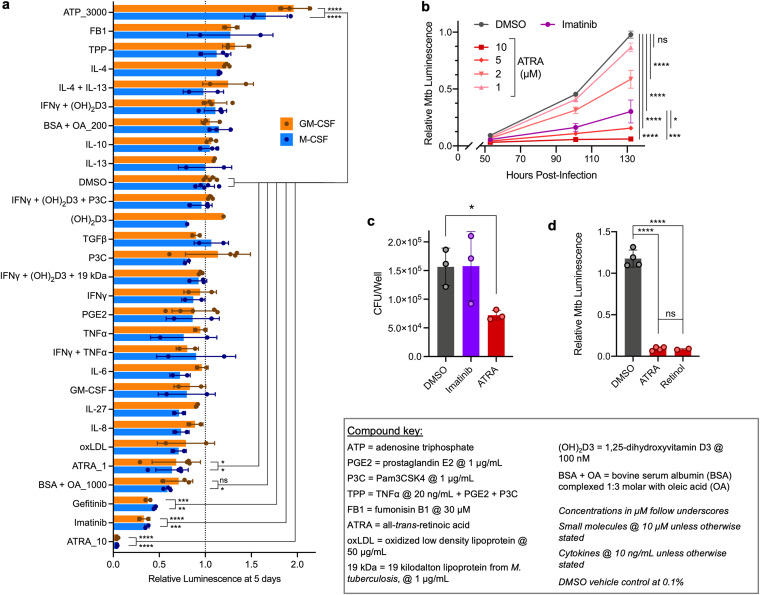
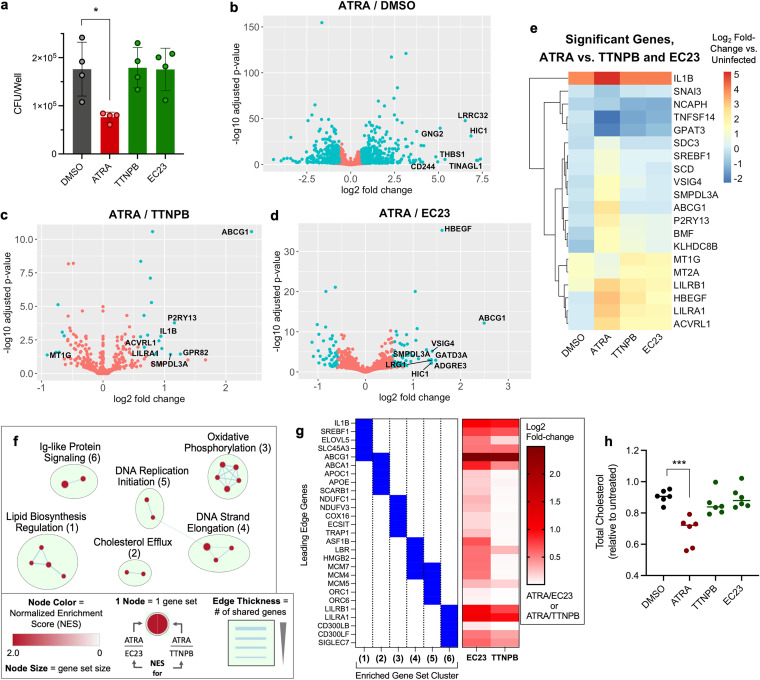
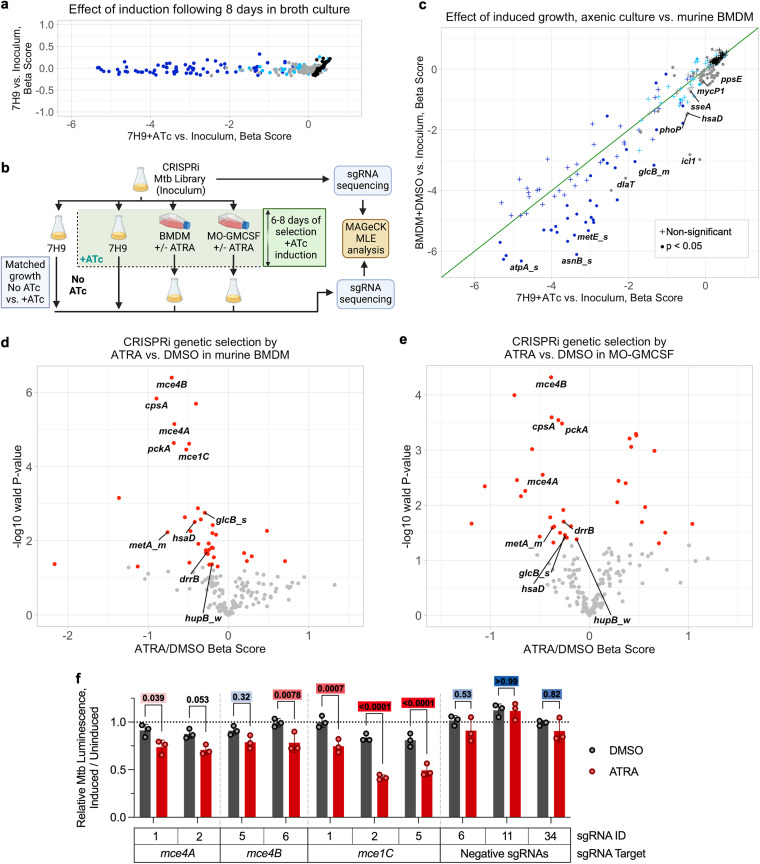
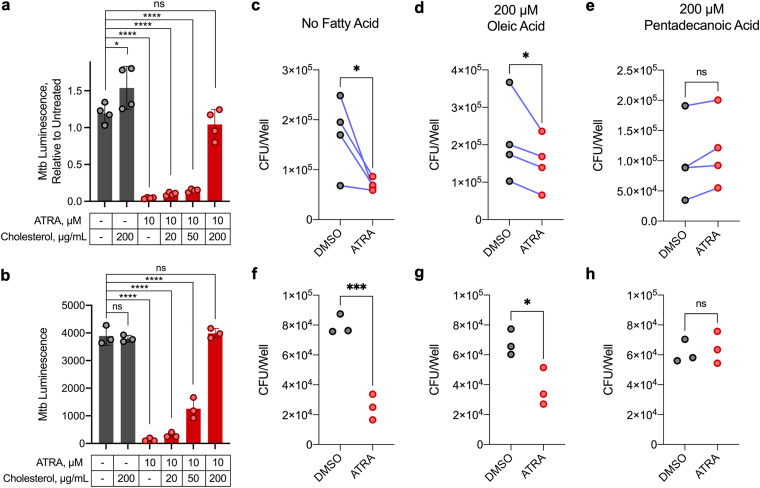
Macrophages are a protective replicative niche for Mycobacterium tuberculosis (Mtb) but can kill the infecting bacterium when appropriately activated. To identify mechanisms of clearance, we compared levels of bacterial restriction by human macrophages after treatment with 26 compounds, including some currently in clinical trials for tuberculosis. All-trans-retinoic acid (ATRA), an active metabolite of vitamin A, drove the greatest increase in Mtb control. Bacterial clearance was transcriptionally and functionally associated with changes in macrophage cholesterol trafficking and lipid metabolism. To determine how these macrophage changes affected bacterial control, we performed the first Mtb CRISPR interference screen in an infection model, identifying Mtb genes specifically required to survive in ATRA-activated macrophages. These data showed that ATRA treatment starves Mtb of cholesterol and the downstream metabolite propionyl coenzyme A (propionyl-CoA). Supplementation with sources of propionyl-CoA, including cholesterol, abrogated the restrictive effect of ATRA. This work demonstrates that targeting the coupled metabolism of Mtb and the macrophage improves control of infection and that it is possible to genetically map the mode of bacterial death using CRISPR interference. IMPORTANCE Tuberculosis, caused by the bacterium Mycobacterium tuberculosis, is a leading cause of death due to infectious disease. Improving the immune response to tuberculosis holds promise for fighting the disease but is limited by our lack of knowledge as to how the immune system kills M. tuberculosis. Our research identifies a potent way to make relevant immune cells more effective at fighting M. tuberculosis and then uses paired human and bacterial genomic methods to determine the mechanism of that improved bacterial clearance.
Keywords: CRISPR interference; Mycobacterium tuberculosis; cholesterol; macrophages; nutritional immunity; propionyl-CoA; retinoic acid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fabri M, Stenger S, Shin D, Yuk J, Liu PT, Realegeno S, Lee H, Krutzik SR, Schenk M, Sieling PA, Teles R, Montoya D, Iyer SS, Bruns H, Lewinsohn DM, Hollis BW, Hewison M, Adams JS, Steinmeyer A, Zügel U, Cheng G, Jo E, Bloom BR, Modlin RL. 2011. Vitamin D is required for IFN-γ-mediated antimicrobial activity of human macrophages. Sci Transl Med 3:104ra102. doi:10.1126/scitranslmed.3003045. - DOI - PMC - PubMed
-
- Bryson BD, Rosebrock TR, Tafesse FG, Itoh CY, Nibasumba A, Babunovic GH, Corleis B, Martin C, Keegan C, Andrade P, Realegeno S, Kwon D, Modlin RL, Fortune SM. 2019. Heterogeneous GM-CSF signaling in macrophages is associated with control of Mycobacterium tuberculosis. Nat Commun 10:2329. doi:10.1038/s41467-019-10065-8. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
