

A neuron-to-astrocyte Wnt5a signal governs astrogliosis during HIV-associated pain pathogenesis
- PMID: 35040478
- PMCID: PMC10200293
- DOI: 10.1093/brain/awac015
A neuron-to-astrocyte Wnt5a signal governs astrogliosis during HIV-associated pain pathogenesis
Abstract
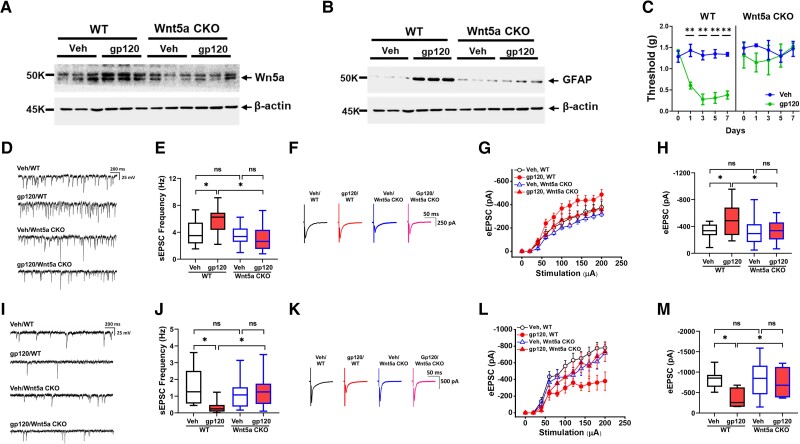
Chronic pain is the most common neurological disorder of HIV patients. Multiple neuropathologies were identified in the pain pathway. Among them is the prominent astrocytic reaction (also know an astrogliosis). However, the pathogenic role and mechanism of the astrogliosis are unclear. Here, we show that the astrogliosis is crucial for the pain development induced by a key neurotoxic HIV protein gp120 and that a neuron-to-astrocyte Wnt5a signal controls the astrogliosis. Ablation of astrogliosis blocked the development of gp120-induced mechanical hyperalgesia, and concomitantly the expression of neural circuit polarization in the spinal dorsal horn. We demonstrated that conditional knockout of either Wnt5a in neurons or its receptor ROR2 in astrocytes abolished not only gp120-induced astrogliosis but also hyperalgesia and neural circuit polarization. Furthermore, we found that the astrogliosis promoted expression of hyperalgesia and NCP via IL-1β regulated by a Wnt5a-ROR2-MMP2 axis. Our results shed light on the role and mechanism of astrogliosis in the pathogenesis of HIV-associated pain.
Keywords: HIV-1 gp120; Wnt; astrocyte; neural circuit; pain.
© The Author(s) 2022. Published by Oxford University Press on behalf of the Guarantors of Brain. All rights reserved. For permissions, please e-mail: journals.permissions@oup.com.
Conflict of interest statement
The authors report no competing interests.
Figures






Similar articles
-
Development of opioid-induced hyperalgesia depends on reactive astrocytes controlled by Wnt5a signaling.Mol Psychiatry. 2023 Feb;28(2):767-779. doi: 10.1038/s41380-022-01815-0. Epub 2022 Oct 6. Mol Psychiatry. 2023. PMID: 36203006 Free PMC article.
-
Wingless-type mammary tumor virus integration site family, member 5A (Wnt5a) regulates human immunodeficiency virus type 1 (HIV-1) envelope glycoprotein 120 (gp120)-induced expression of pro-inflammatory cytokines via the Ca2+/calmodulin-dependent protein kinase II (CaMKII) and c-Jun N-terminal kinase (JNK) signaling pathways.J Biol Chem. 2013 May 10;288(19):13610-9. doi: 10.1074/jbc.M112.381046. Epub 2013 Mar 28. J Biol Chem. 2013. PMID: 23539626 Free PMC article.
-
Microglia Mediate HIV-1 gp120-Induced Synaptic Degeneration in Spinal Pain Neural Circuits.J Neurosci. 2019 Oct 16;39(42):8408-8421. doi: 10.1523/JNEUROSCI.2851-18.2019. Epub 2019 Aug 30. J Neurosci. 2019. PMID: 31471472 Free PMC article.
-
Neural Circuitry Polarization in the Spinal Dorsal Horn (SDH): A Novel Form of Dysregulated Circuitry Plasticity during Pain Pathogenesis.Cells. 2024 Feb 25;13(5):398. doi: 10.3390/cells13050398. Cells. 2024. PMID: 38474361 Free PMC article. Review.
-
Astrocyte activation and apoptosis: their roles in the neuropathology of HIV infection.Brain Pathol. 2003 Jan;13(1):84-94. doi: 10.1111/j.1750-3639.2003.tb00009.x. Brain Pathol. 2003. PMID: 12580548 Free PMC article. Review.
Cited by
-
Wnt signaling pathway in spinal cord injury: from mechanisms to potential applications.Front Mol Neurosci. 2024 Jul 24;17:1427054. doi: 10.3389/fnmol.2024.1427054. eCollection 2024. Front Mol Neurosci. 2024. PMID: 39114641 Free PMC article. Review.
-
GPR37L1 identifies spinal cord astrocytes and protects neuropathic pain after nerve injury.Neuron. 2025 Apr 16;113(8):1206-1222.e6. doi: 10.1016/j.neuron.2025.01.012. Epub 2025 Feb 13. Neuron. 2025. PMID: 39952243
-
Hevin/Sparcl1 drives pathological pain through spinal cord astrocyte and NMDA receptor signaling.JCI Insight. 2022 Dec 8;7(23):e161028. doi: 10.1172/jci.insight.161028. JCI Insight. 2022. PMID: 36256481 Free PMC article.
-
Provoked cytokine response is not associated with distress or induced secondary hyperalgesia in people with suppressed HIV.medRxiv [Preprint]. 2025 May 27:2025.01.21.25320673. doi: 10.1101/2025.01.21.25320673. medRxiv. 2025. Update in: Pain. 2025 Aug 7. doi: 10.1097/j.pain.0000000000003748. PMID: 39973982 Free PMC article. Updated. Preprint.
-
Astrocytes in Chronic Pain: Cellular and Molecular Mechanisms.Neurosci Bull. 2023 Mar;39(3):425-439. doi: 10.1007/s12264-022-00961-3. Epub 2022 Nov 14. Neurosci Bull. 2023. PMID: 36376699 Free PMC article. Review.
References
-
- Sabin CA, Harding R, Bagkeris E, et al. . Pain in people living with HIV and its association with healthcare resource use, well being and functional status. AIDS. 2018;32(18):2697–2706. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
