

Impaired Oligodendrocyte Maturation Is an Early Feature in SCA3 Disease Pathogenesis
- PMID: 35042771
- PMCID: PMC8883861
- DOI: 10.1523/JNEUROSCI.1954-20.2021
Impaired Oligodendrocyte Maturation Is an Early Feature in SCA3 Disease Pathogenesis
Abstract
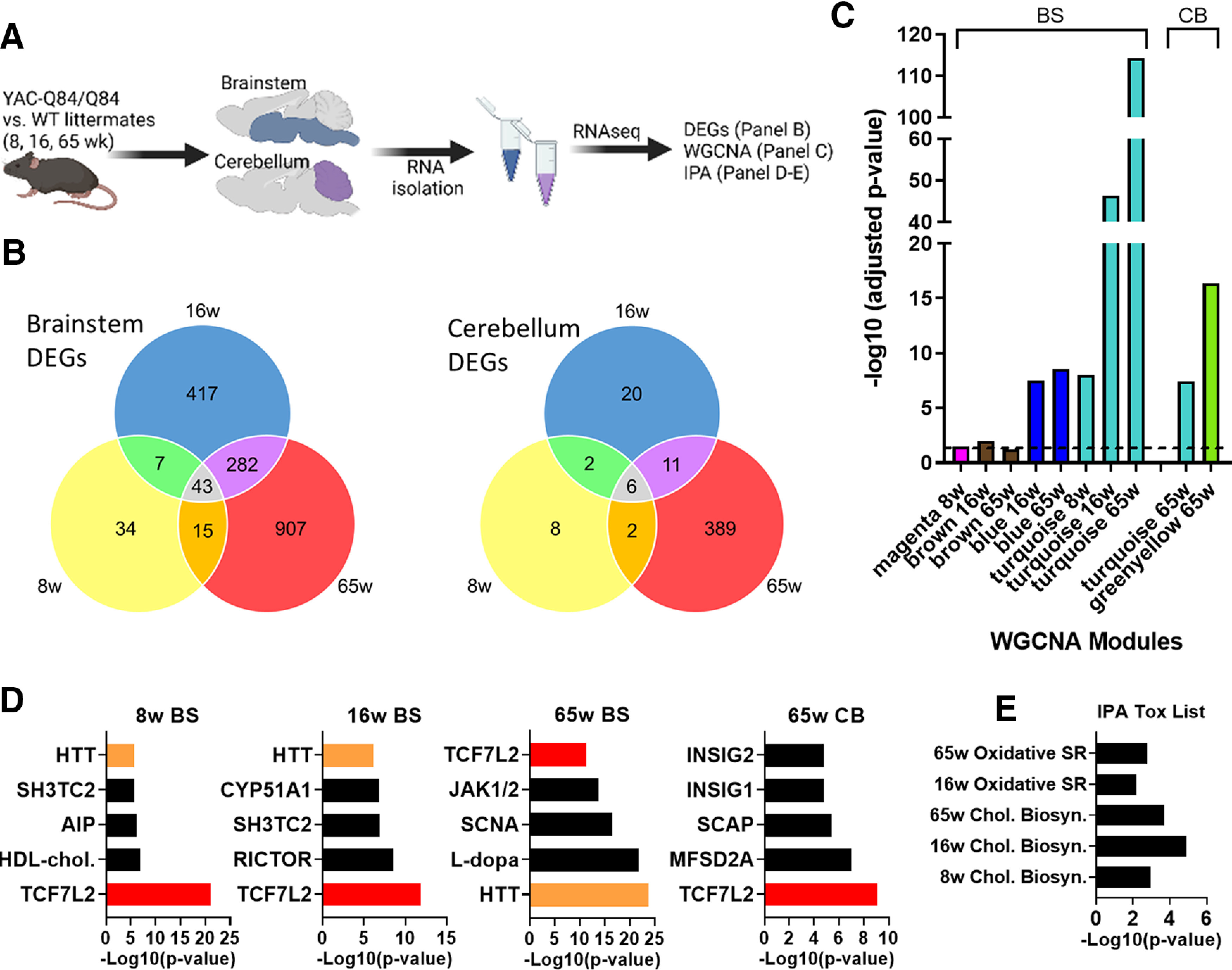
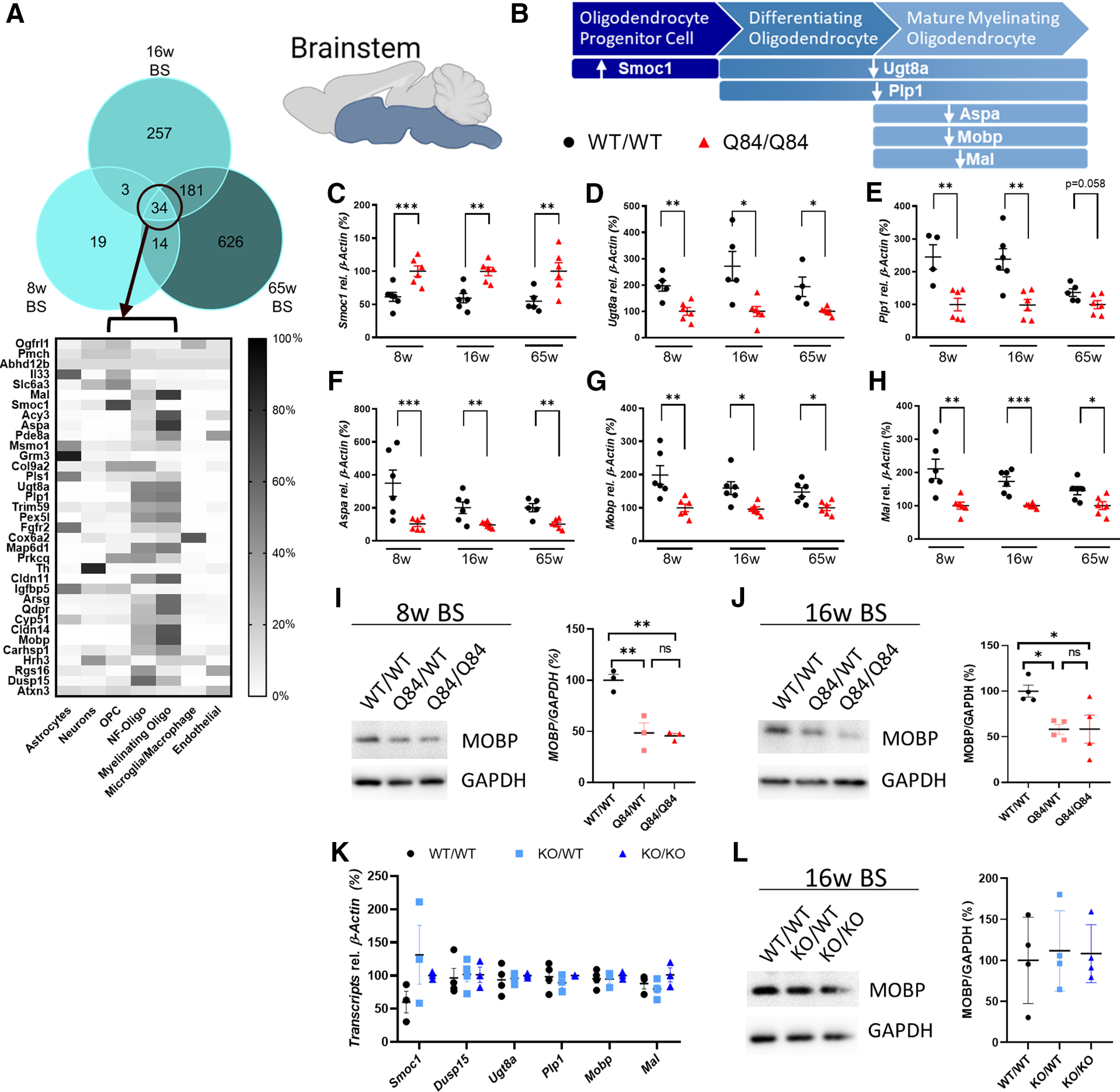
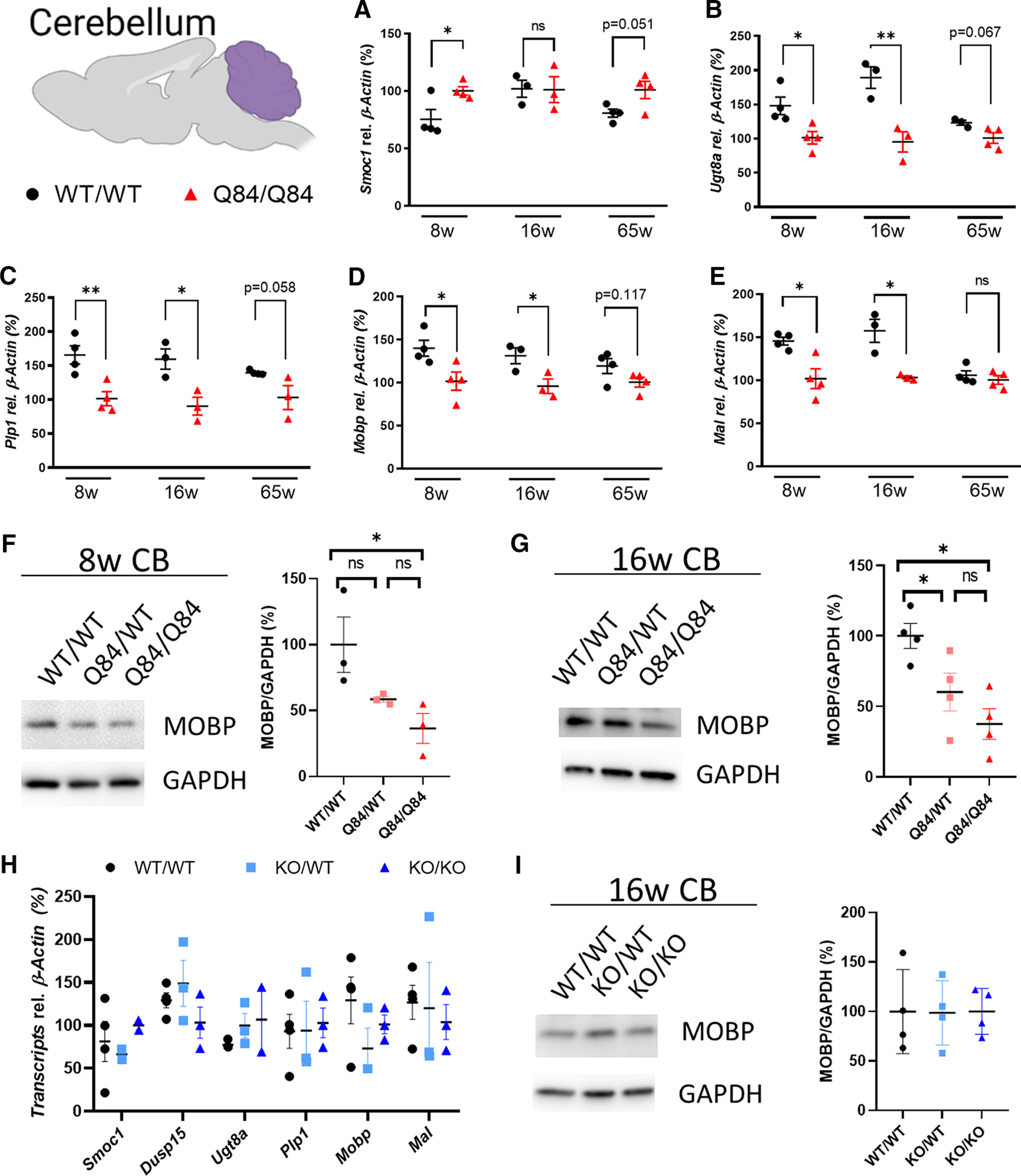
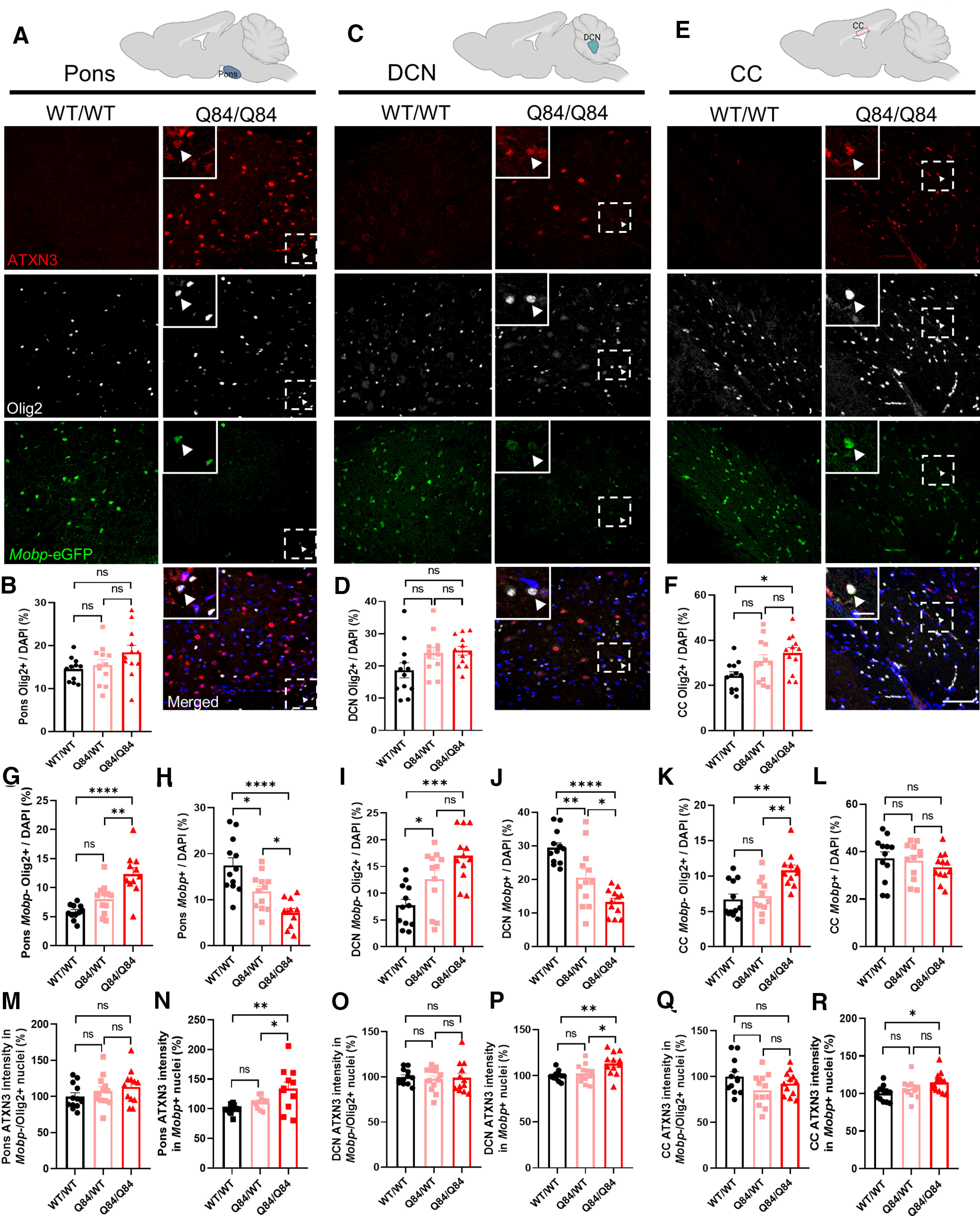
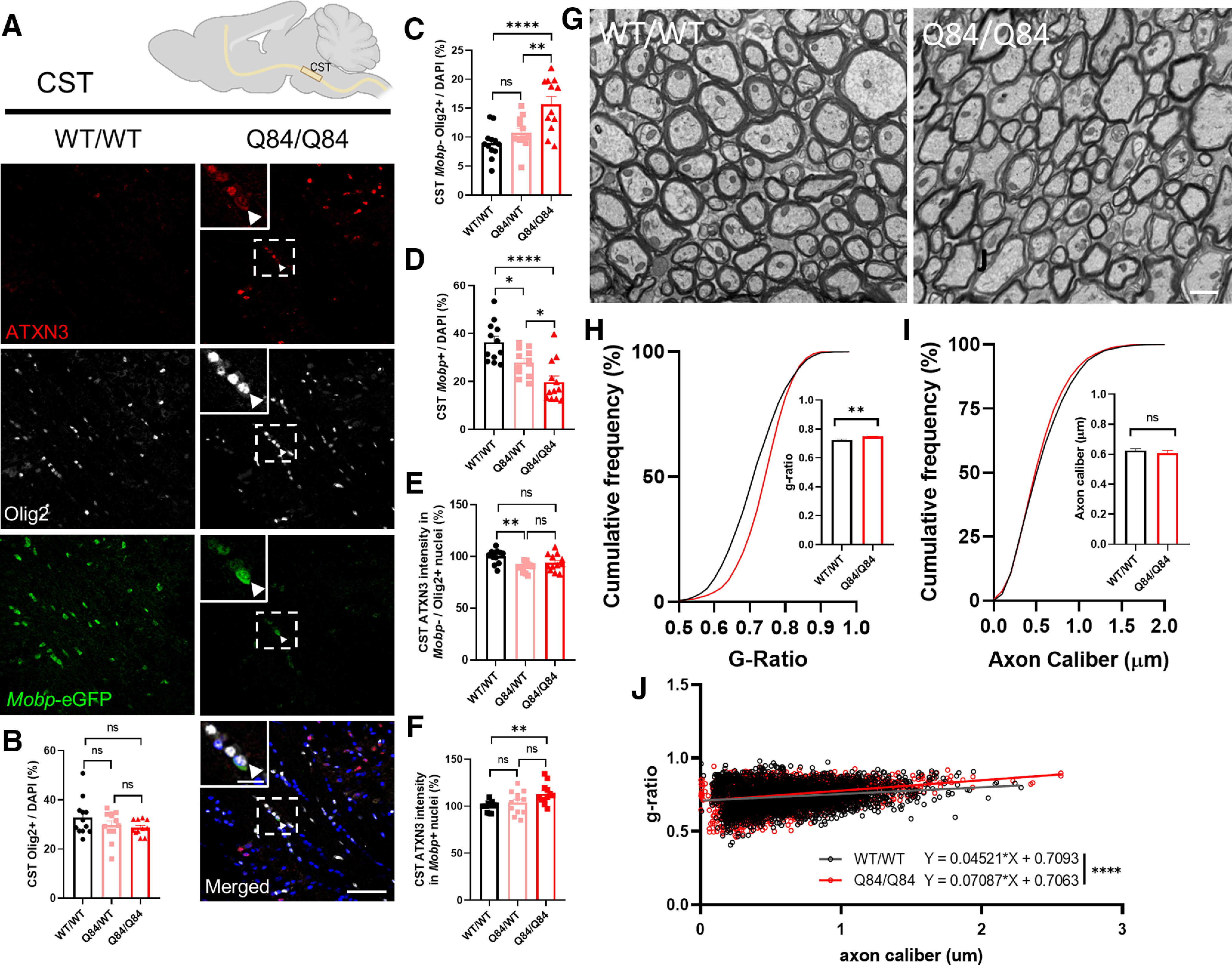
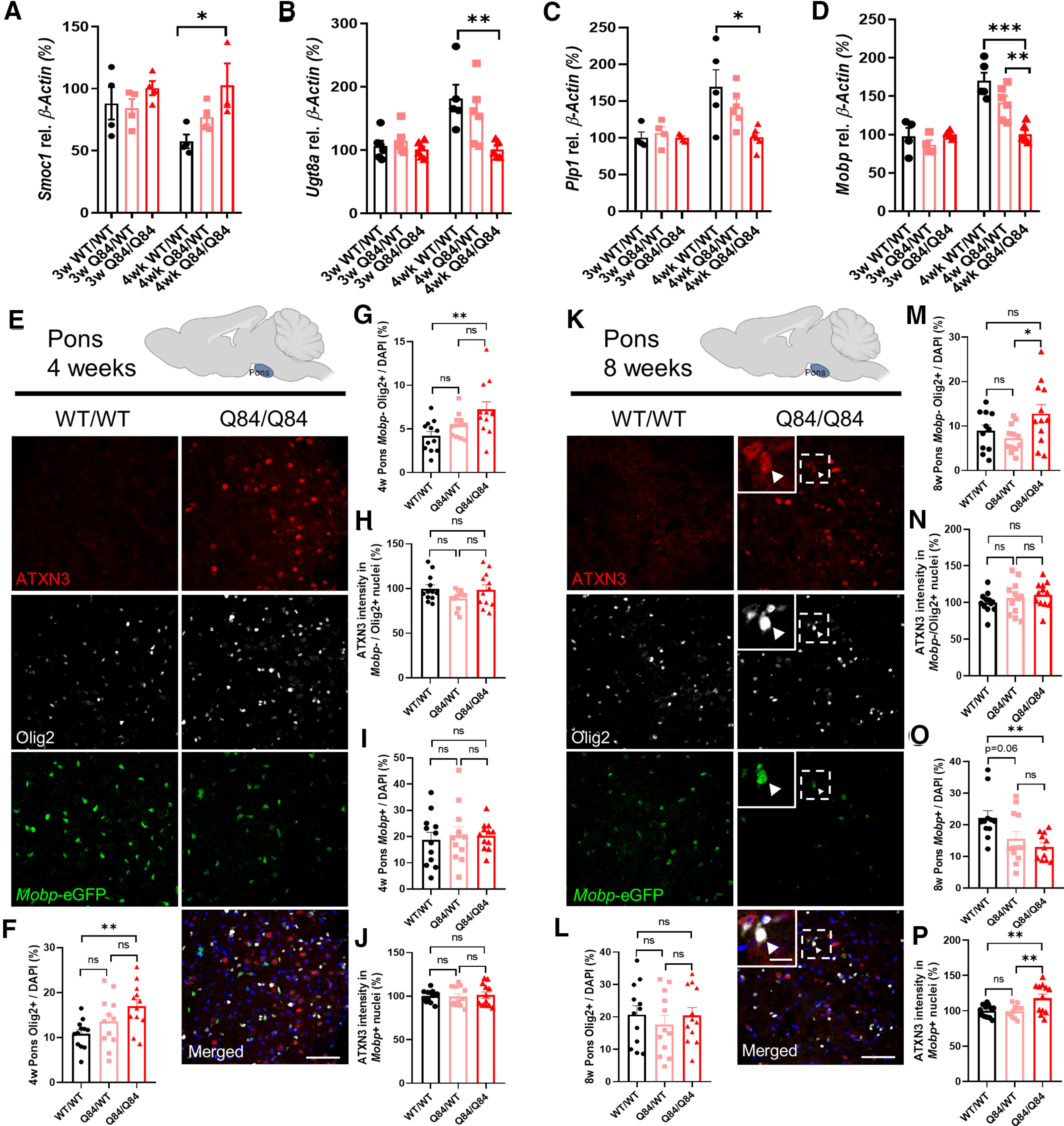
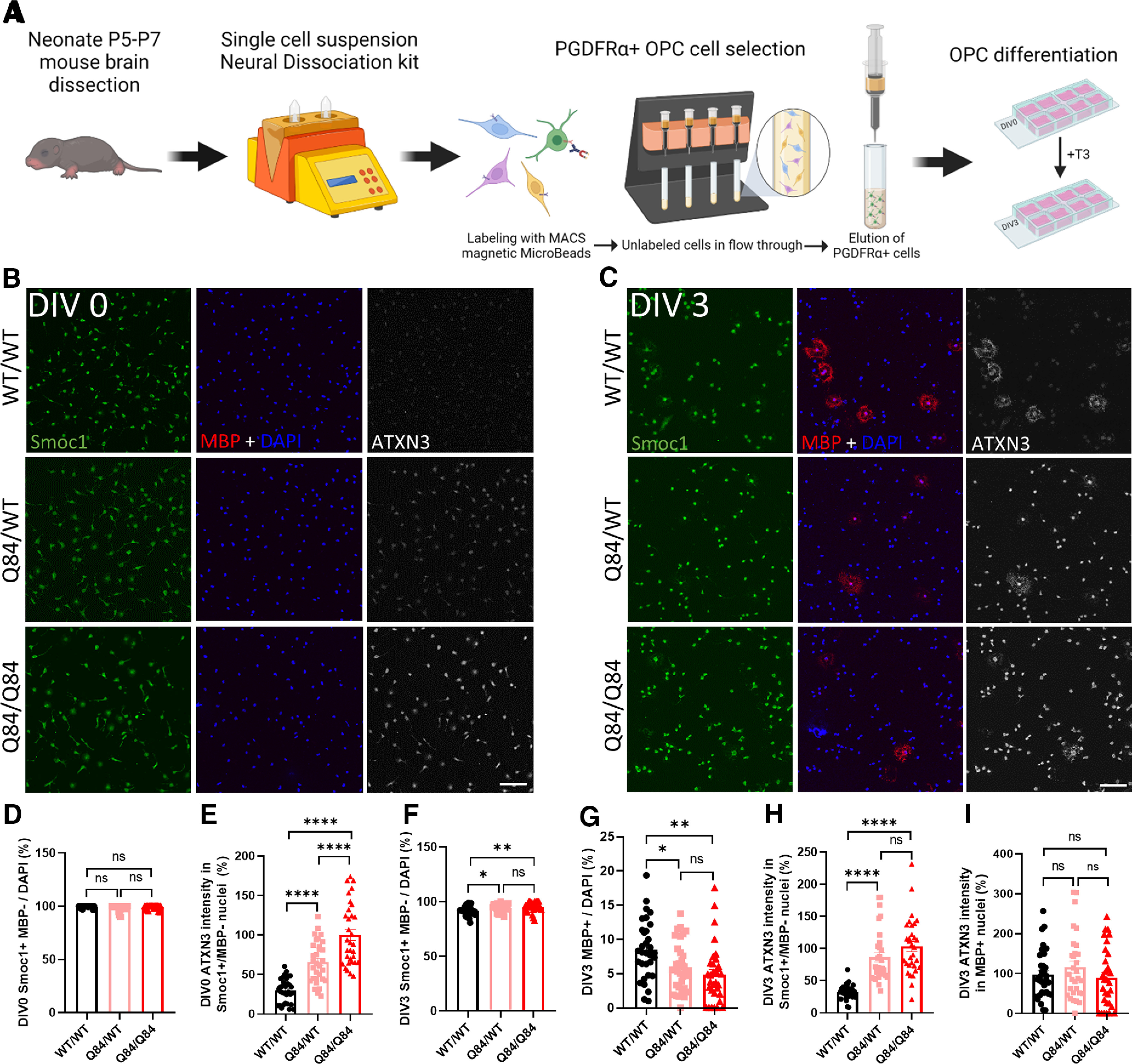
Spinocerebellar ataxia Type 3 (SCA3), the most common dominantly inherited ataxia, is a polyglutamine neurodegenerative disease for which there is no disease-modifying therapy. The polyglutamine-encoding CAG repeat expansion in the ATXN3 gene results in expression of a mutant form of the ATXN3 protein, a deubiquitinase that causes selective neurodegeneration despite being widely expressed. The mechanisms driving neurodegeneration in SCA3 are unclear. Research to date, however, has focused almost exclusively on neurons. Here, using equal male and female age-matched transgenic mice expressing full-length human mutant ATXN3, we identified early and robust transcriptional changes in selectively vulnerable brain regions that implicate oligodendrocytes in disease pathogenesis. We mapped transcriptional changes across early, mid, and late stages of disease in two selectively vulnerable brain regions: the cerebellum and brainstem. The most significant disease-associated module through weighted gene coexpression network analysis revealed dysfunction in SCA3 oligodendrocyte maturation. These results reflect a toxic gain-of-function mechanism, as ATXN3 KO mice do not exhibit any impairments in oligodendrocyte maturation. Genetic crosses to reporter mice revealed a marked reduction in mature oligodendrocytes in SCA3-disease vulnerable brain regions, and ultrastructural microscopy confirmed abnormalities in axonal myelination. Further study of isolated oligodendrocyte precursor cells from SCA3 mice established that this impairment in oligodendrocyte maturation is a cell-autonomous process. We conclude that SCA3 is not simply a disease of neurons, and the search for therapeutic strategies and disease biomarkers will need to account for non-neuronal involvement in SCA3 pathogenesis.SIGNIFICANCE STATEMENT Despite advances in spinocerebellar ataxia Type 3 (SCA3) disease understanding, much remains unknown about how the disease gene causes brain dysfunction ultimately leading to cell death. We completed a longitudinal transcriptomic analysis of vulnerable brain regions in SCA3 mice to define the earliest and most robust changes across disease progression. Through gene network analyses followed up with biochemical and histologic studies in SCA3 mice, we provide evidence for severe dysfunction in oligodendrocyte maturation early in SCA3 pathogenesis. Our results advance understanding of SCA3 disease mechanisms, identify additional routes for therapeutic intervention, and may provide broader insight into polyglutamine diseases beyond SCA3.
Keywords: Machado–Joseph disease; ataxia; myelination; oligodendrocyte; polyglutamine; spinocerebellar ataxia Type 3.
Copyright © 2022 the authors.
Figures
References
-
- Bichelmeier U, Schmidt T, Hübener J, Boy J, Rüttiger L, Häbig K, Poths S, Bonin M, Knipper M, Schmidt WJ, Wilbertz J, Wolburg H, Laccone F, Riess O (2007) Nuclear localization of Ataxin-3 is required for the manifestation of symptoms in SCA3: in vivo evidence. J Neurosci 27:7418–7428. 10.1523/JNEUROSCI.4540-06.2007 - DOI - PMC - PubMed
-
- Cemal CK, Carroll CJ, Lawrence L, Lowrie MB, Ruddle P, Al-Mahdawi S, King RH, Pook MA, Huxley C, Chamberlain S (2002) YAC transgenic mice carrying pathological alleles of the MJD1 locus exhibit a mild and slowly progressive cerebellar deficit. Hum Mol Genet 11:1075–1094. 10.1093/hmg/11.9.1075 - DOI - PubMed
-
- do Carmo Costa M, Luna-Cancalon K, Fischer S, Ashraf NS, Ouyang M, Dharia RM, Martin-Fishman L, Yang Y, Shakkottai VG, Davidson BL, Rodríguez-Lebrón E, Paulson HL (2013) Toward RNAi therapy for the polyglutamine disease Machado–Joseph disease. Mol Ther 21:1898–1908. 10.1038/mt.2013.144 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials