

PROTAC targeted protein degraders: the past is prologue
- PMID: 35042991
- PMCID: PMC8765495
- DOI: 10.1038/s41573-021-00371-6
PROTAC targeted protein degraders: the past is prologue
Abstract
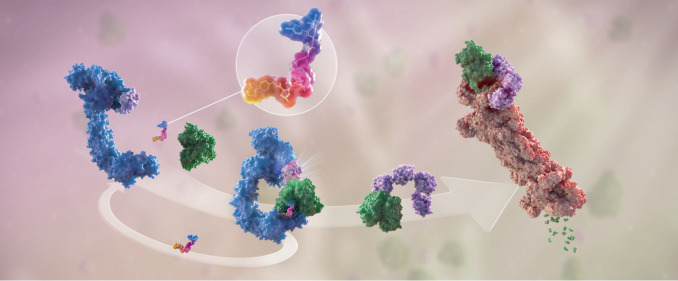
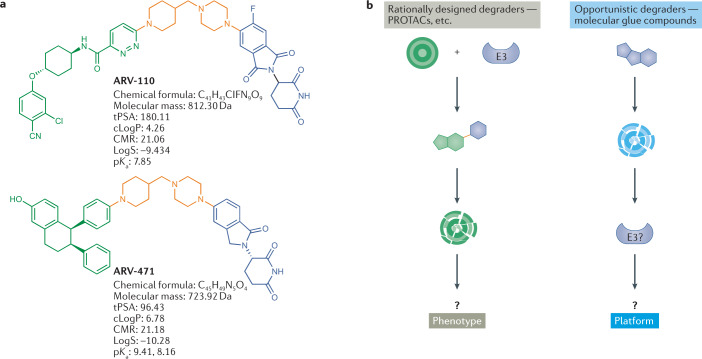
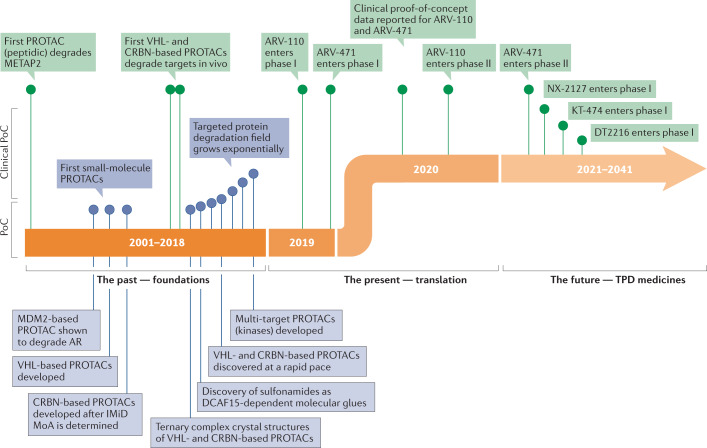
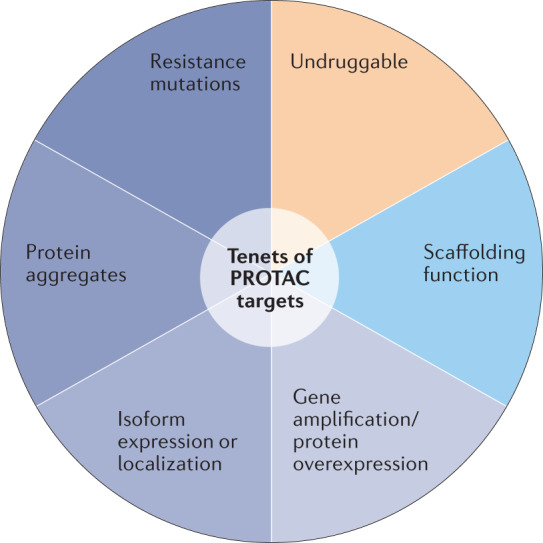
Targeted protein degradation (TPD) is an emerging therapeutic modality with the potential to tackle disease-causing proteins that have historically been highly challenging to target with conventional small molecules. In the 20 years since the concept of a proteolysis-targeting chimera (PROTAC) molecule harnessing the ubiquitin-proteasome system to degrade a target protein was reported, TPD has moved from academia to industry, where numerous companies have disclosed programmes in preclinical and early clinical development. With clinical proof-of-concept for PROTAC molecules against two well-established cancer targets provided in 2020, the field is poised to pursue targets that were previously considered 'undruggable'. In this Review, we summarize the first two decades of PROTAC discovery and assess the current landscape, with a focus on industry activity. We then discuss key areas for the future of TPD, including establishing the target classes for which TPD is most suitable, expanding the use of ubiquitin ligases to enable precision medicine and extending the modality beyond oncology.
© 2022. Springer Nature Limited.
Conflict of interest statement
C.M.C. is a consultant and shareholder in Arvinas, Inc. and Halda Therapeutics, which support research in his lab. M.B. and D.R.L. are employees and shareholders of Arvinas, Inc, which is developing drug candidates in the targeted protein degradation space.
Figures
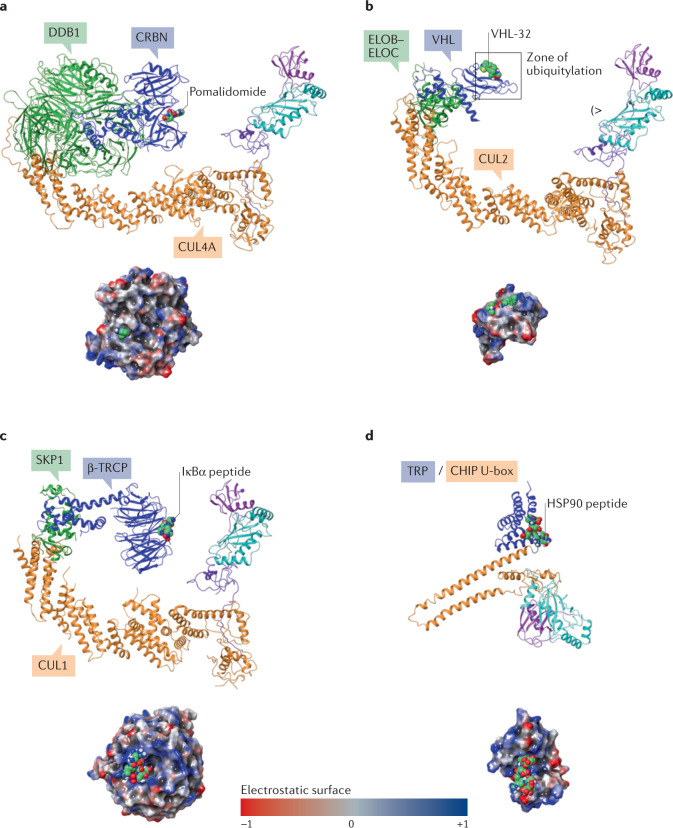
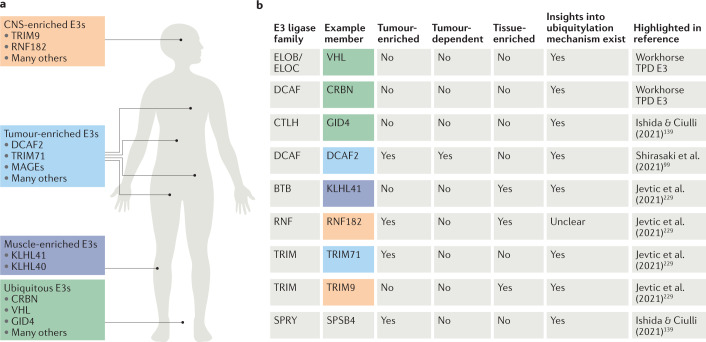
References
-
- Hansen JD, et al. Discovery of CRBN E3 ligase modulator CC-92480 for the treatment of relapsed and refractory multiple myeloma. J. Med. Chem. 2020;63:6648–6676. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
