

This is a preprint.
Shift of lung macrophage composition is associated with COVID-19 disease severity and recovery
- PMID: 35043110
- PMCID: PMC8764718
- DOI: 10.1101/2022.01.11.475918
Shift of lung macrophage composition is associated with COVID-19 disease severity and recovery
Update in
-
A shift in lung macrophage composition is associated with COVID-19 severity and recovery.Sci Transl Med. 2022 Sep 14;14(662):eabn5168. doi: 10.1126/scitranslmed.abn5168. Epub 2022 Sep 14. Sci Transl Med. 2022. PMID: 36103512 Free PMC article.
Abstract
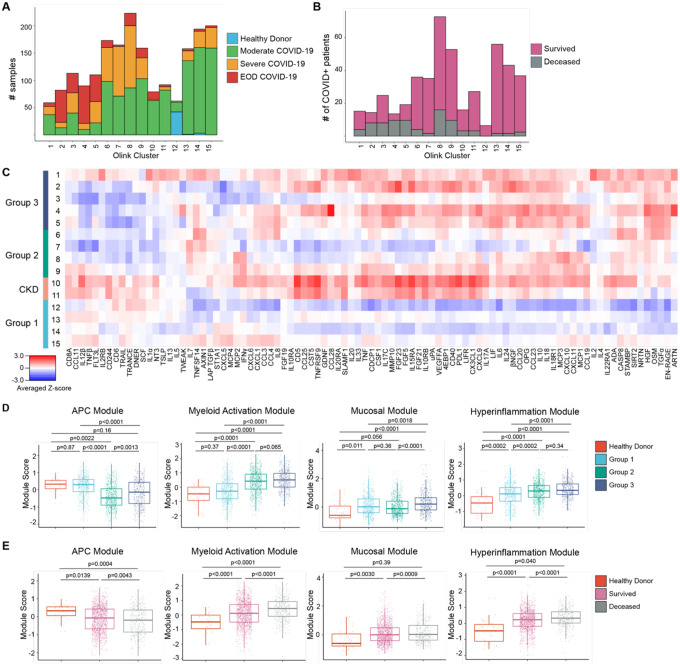
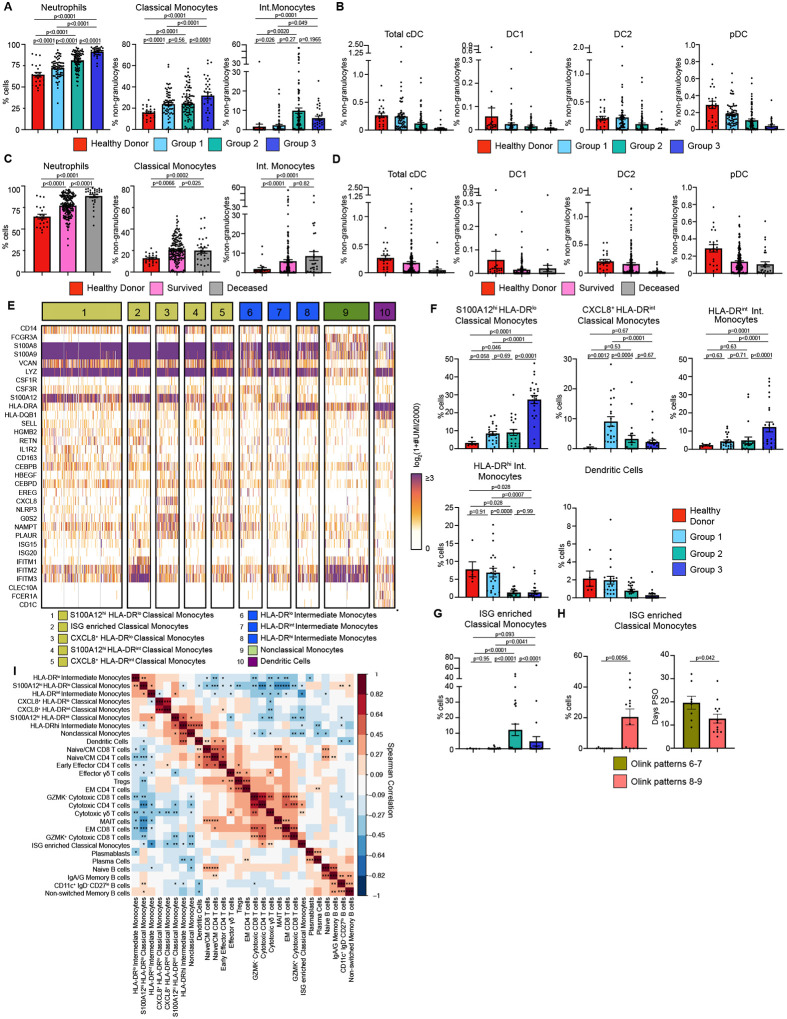
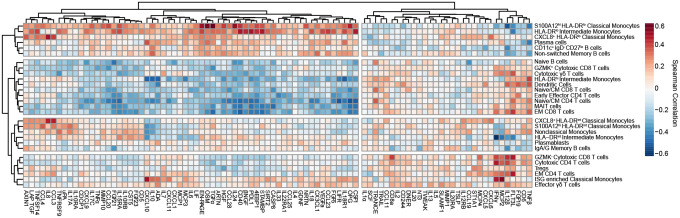
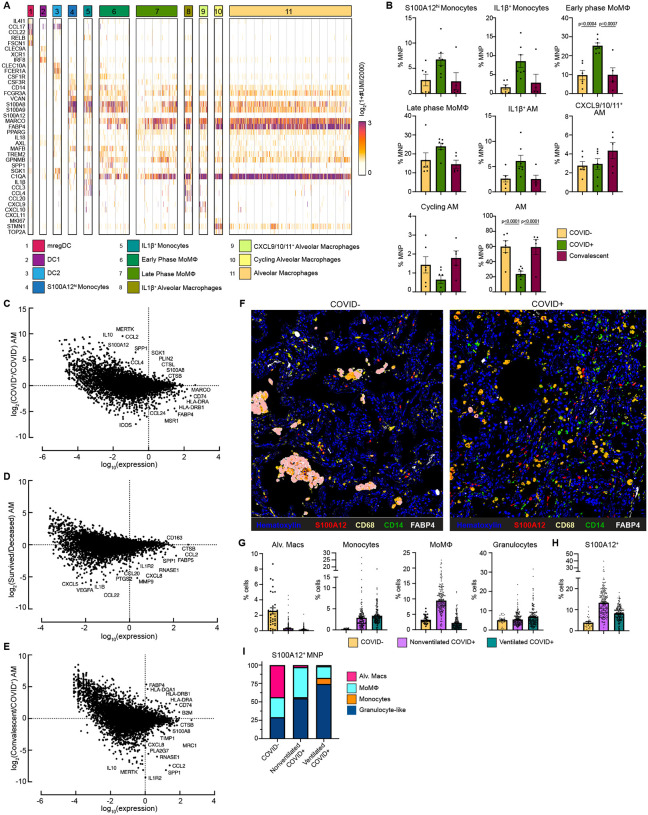
Though it has been 2 years since the start of the Coronavirus Disease 19 (COVID-19) pandemic, COVID-19 continues to be a worldwide health crisis. Despite the development of preventive vaccines, very little progress has been made to identify curative therapies to treat COVID-19 and other inflammatory diseases which remain a major unmet need in medicine. Our study sought to identify drivers of disease severity and death to develop tailored immunotherapy strategies to halt disease progression. Here we assembled the Mount Sinai COVID-19 Biobank which was comprised of ~600 hospitalized patients followed longitudinally during the peak of the pandemic. Moderate disease and survival were associated with a stronger antigen (Ag) presentation and effector T cell signature, while severe disease and death were associated with an altered Ag presentation signature, increased numbers of circulating inflammatory, immature myeloid cells, and extrafollicular activated B cells associated with autoantibody formation. Strikingly, we found that in severe COVID-19 patients, lung tissue resident alveolar macrophages (AM) were not only severely depleted, but also had an altered Ag presentation signature, and were replaced by inflammatory monocytes and monocyte-derived macrophages (MoMΦ). Notably, the size of the AM pool correlated with recovery or death, while AM loss and functionality were restored in patients that recovered. These data therefore suggest that local and systemic myeloid cell dysregulation is a driver of COVID-19 severity and that modulation of AM numbers and functionality in the lung may be a viable therapeutic strategy for the treatment of critical lung inflammatory illnesses.
Figures
References
-
- Charney A. W., Simons N. W., Mouskas K., Lepow L., Cheng E., Le Berichel J., Chang C., Marvin R., Del Valle D. M., Calorossi S., Lansky A., Walker L., Patel M., Xie H., Yi N., Yu A., Kang G., Liharska L. E., Moya E., Hartnett M., Hatem S., Wilkins L., Eaton M., Jamal H., Tuballes K., Chen S. T., Chung J., Harris J., Batchelor C., Lacunza J., Yishak M., Argueta K., Karekar N., Lee B., Kelly G., Geanon D., Handler D., Leech J., Stefanos H., Dawson T., Scott I., Francoeur N., Johnson J. S., Vaid A., Glicksberg B. S., Nadkarni G. N., Schadt E. E., Gelb B. D., Rahman A., Sebra R., Martin G., Agashe C., Agrawal P., Alesso-Carra K., Alibo E., Alvarez K., Amabile A., Ascolillo S., Bailey R., Begani P., Correra P. B., Brown S. A., Buckup M., Burka L., Cambron L., Carrara G., Chang S., Chien J., Chowdhury M., Bozkus C. C., Comella P., Cosgrove D., Cossarini F., Cotter L., Dave A., Dayal B., Dhainaut M., Dornfeld R., Dul K., Eber N., Elaiho C., Fabris F., Faith J., Falci D., Feng S., Fennessy B., Fernandes M., Gangadharan S., Grabowska J., Grimes G., Hamdani M., Herbinet M., Herrera E., Hochman A., Hoffman G. E., Hook J., Horta L., Humblin E., Karim S., Kim J., Lebovitch D., Lee G., Lee G. H., Lee J., Leventhal M., Lindblad K., Livanos A., Machado R., Mahmood Z., Mar K., Maskey S., Matthews P., Meckel K., Mehandru S., Mercedes C., Meyer D., Mollaoglu G., Morris S., Nie K., Nisenholtz M., Ofori-Amanfo G., Onel K., Ounadjela M., Patel V., Pruitt C., Rathi S., Redes J., Reyes-Torres I., Rodrigues A., Rodriguez A., Roudko V., Ruiz E., Scalzo P., Silva P., Schanoski A. S., Straw M., Tabachnikova S., Teague C., Upadhyaya B., Van Der Heide V., Vaninov N., Wacker D., Walsh H., Wilk C. M., Wilson J., Wilson K. M., Xue L., akomaah Yeboah N., Young S., Zaks N., Zha R., Marron T., Beckmann N., Kim-Schulze S., Gnjatic S., Merad M., Sampling the host response to SARS-CoV-2 in hospitals under siege, Nat. Med. 2020 268 26, 1157–1158 (2020). - PubMed
-
- Locksley R. M., Killeen N., Lenardo M. J., The TNF and TNF Receptor Superfamilies: Integrating Mammalian Biology, Cell 104, 487–501 (2001). - PubMed
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources