

CRLF3 plays a key role in the final stage of platelet genesis and is a potential therapeutic target for thrombocythemia
- PMID: 35051265
- PMCID: PMC7614665
- DOI: 10.1182/blood.2021013113
CRLF3 plays a key role in the final stage of platelet genesis and is a potential therapeutic target for thrombocythemia
Abstract
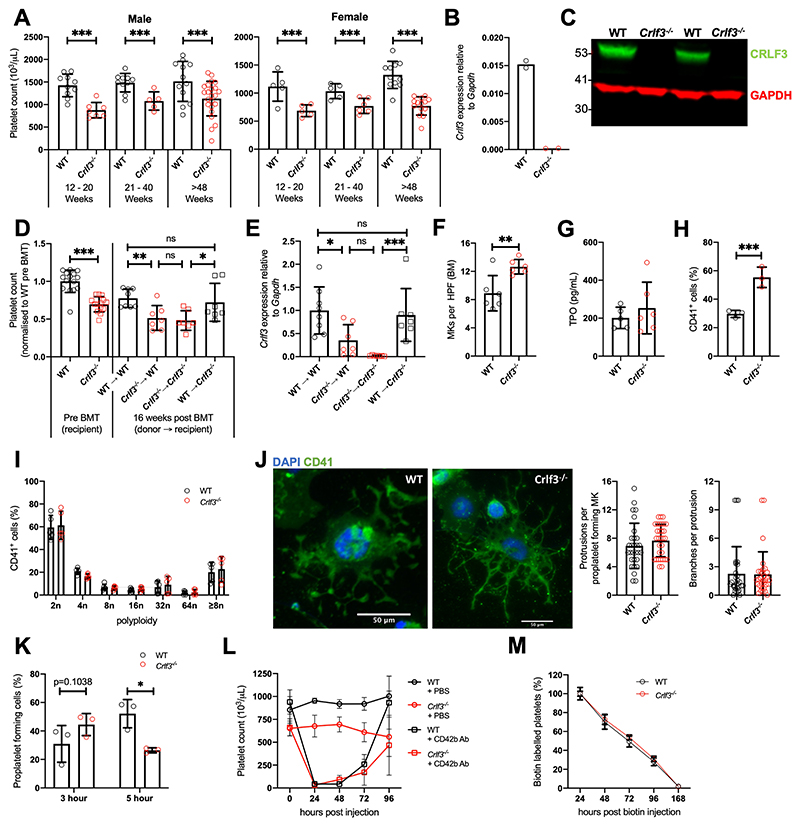
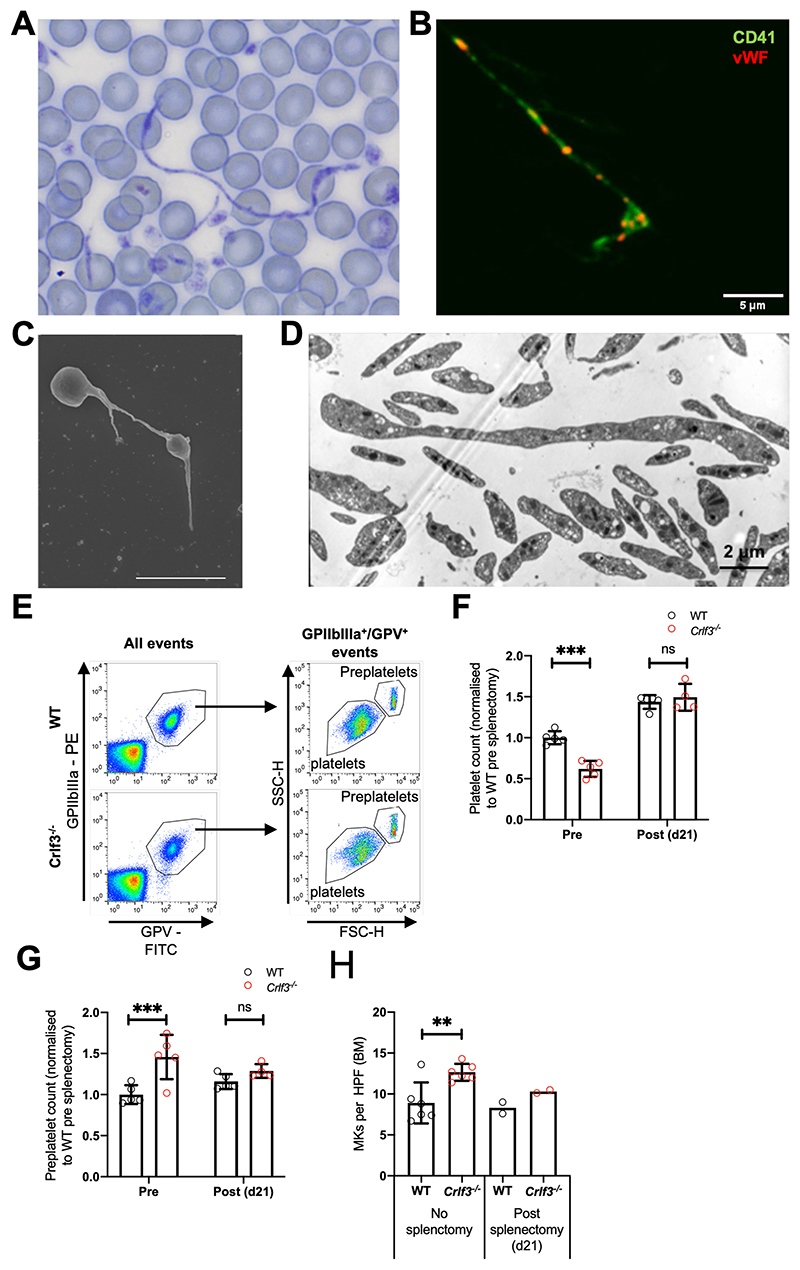
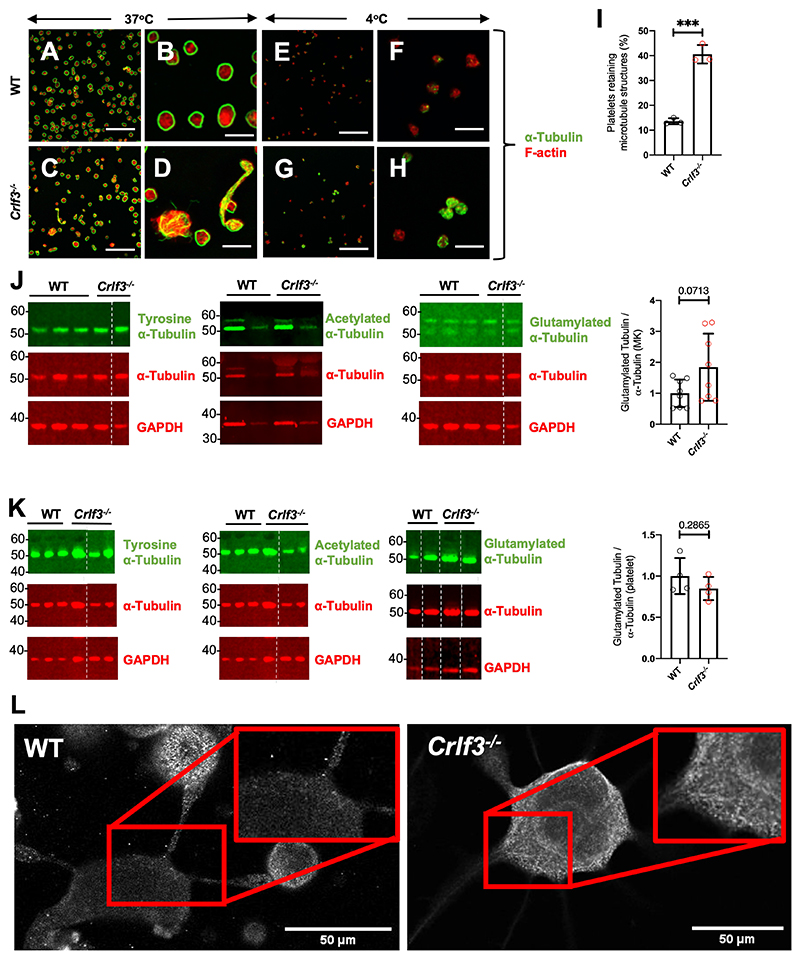
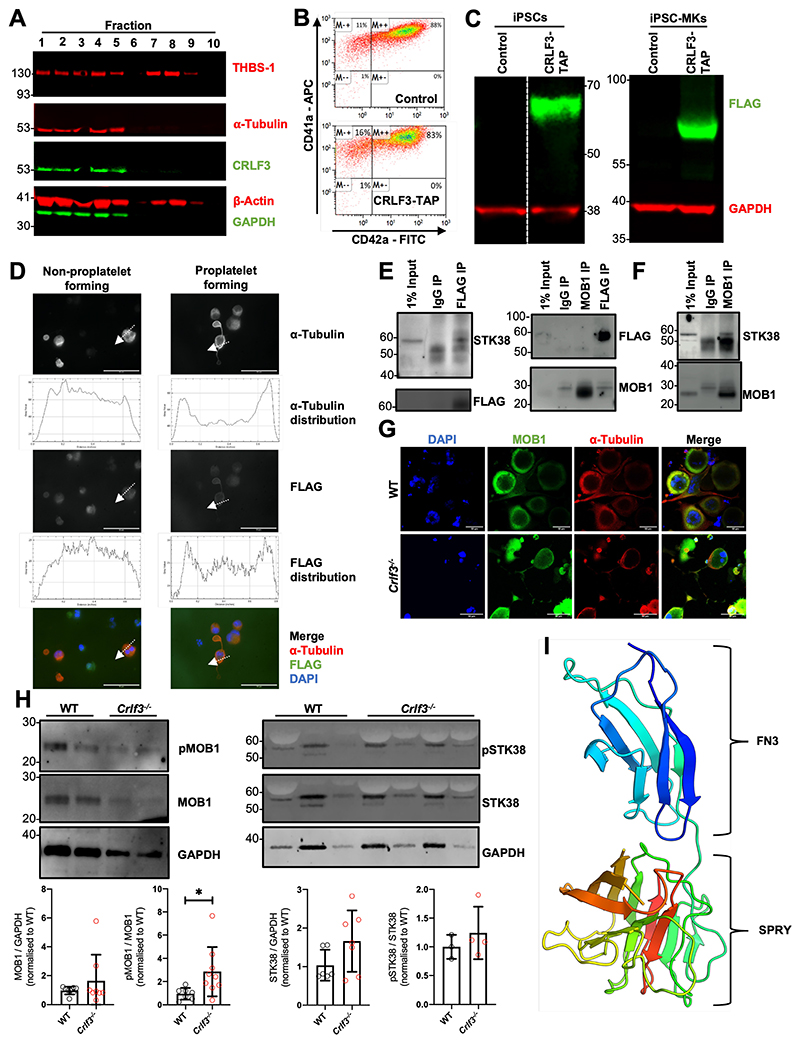
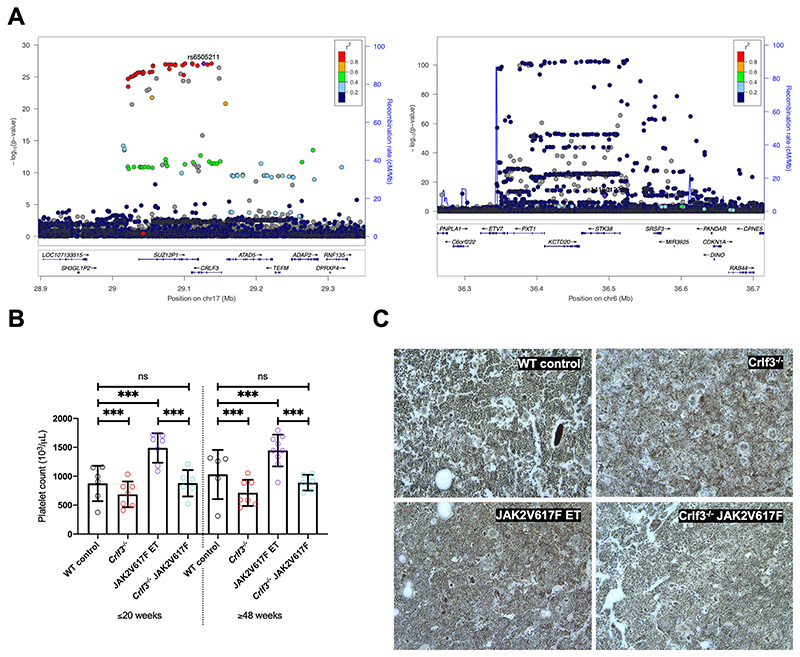
The process of platelet production has so far been understood to be a 2-stage process: megakaryocyte maturation from hematopoietic stem cells followed by proplatelet formation, with each phase regulating the peripheral blood platelet count. Proplatelet formation releases into the bloodstream beads-on-a-string preplatelets, which undergo fission into mature platelets. For the first time, we show that preplatelet maturation is a third, tightly regulated, critical process akin to cytokinesis that regulates platelet count. We show that deficiency in cytokine receptor-like factor 3 (CRLF3) in mice leads to an isolated and sustained 25% to 48% reduction in the platelet count without any effect on other blood cell lineages. We show that Crlf3-/- preplatelets have increased microtubule stability, possibly because of increased microtubule glutamylation via the interaction of CRLF3 with key members of the Hippo pathway. Using a mouse model of JAK2 V617F essential thrombocythemia, we show that a lack of CRLF3 leads to long-term lineage-specific normalization of the platelet count. We thereby postulate that targeting CRLF3 has therapeutic potential for treatment of thrombocythemia.
© 2022 by The American Society of Hematology. Licensed under Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0), permitting only noncommercial, nonderivative use with attribution. All other rights reserved.
Conflict of interest statement
Figures
Comment in
-
The triple crown of platelet generation.Blood. 2022 Apr 7;139(14):2100-2101. doi: 10.1182/blood.2021015018. Blood. 2022. PMID: 35389441 Free PMC article. No abstract available.
References
-
- Junt T, Schulze H, Chen Z, et al. Dynamic visualization of thrombopoiesis within bone marrow. Science. 2007;317(5845):1767–1770. - PubMed
-
- Kunishima S, Nishimura S, Suzuki H, Imaizumi M, Saito H. TUBB1 mutation disrupting microtubule assembly impairs proplatelet formation and results in congenital macrothrombocytopenia. Eur J Haematol. 2014;92(4):276–282. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
