

Evolutionary Origin of Insulin-Degrading Enzyme and Its Subcellular Localization and Secretion Mechanism: A Study in Microglial Cells
- PMID: 35053342
- PMCID: PMC8774118
- DOI: 10.3390/cells11020227
Evolutionary Origin of Insulin-Degrading Enzyme and Its Subcellular Localization and Secretion Mechanism: A Study in Microglial Cells
Abstract
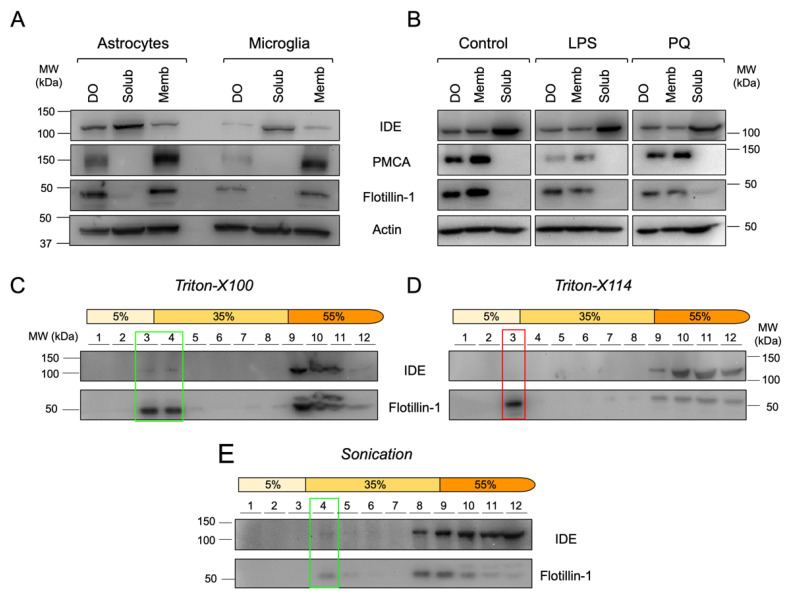
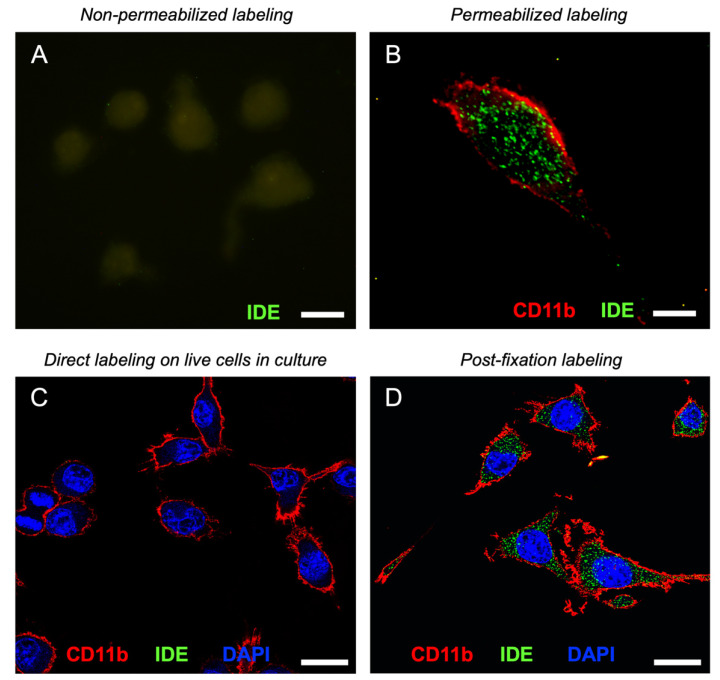
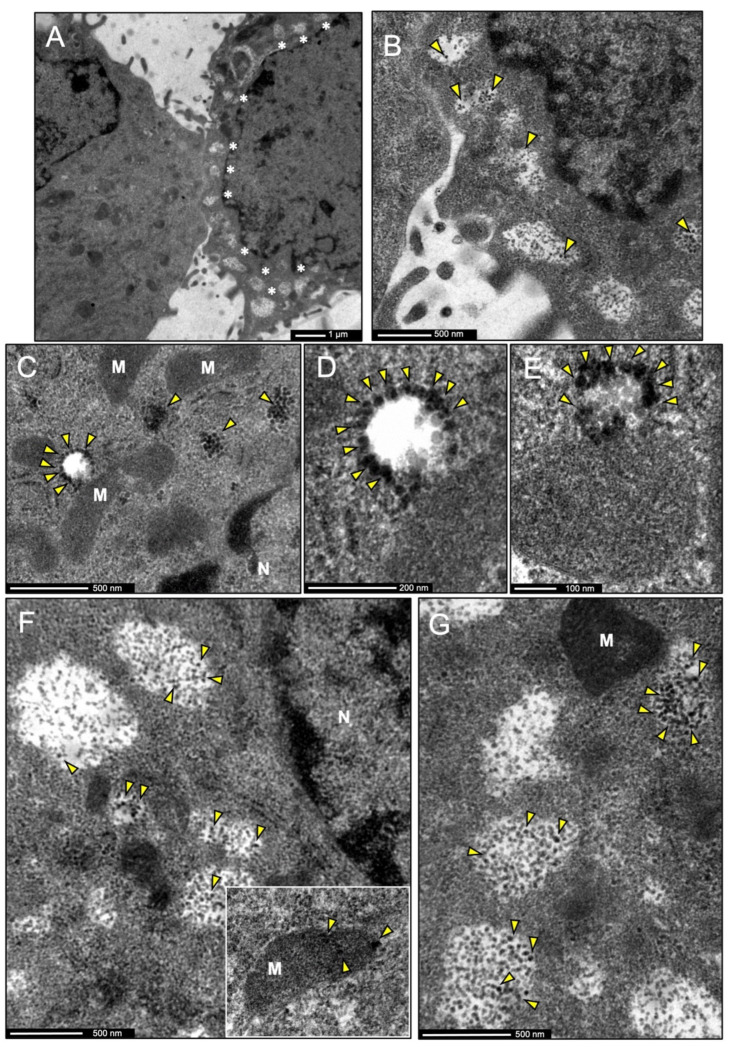
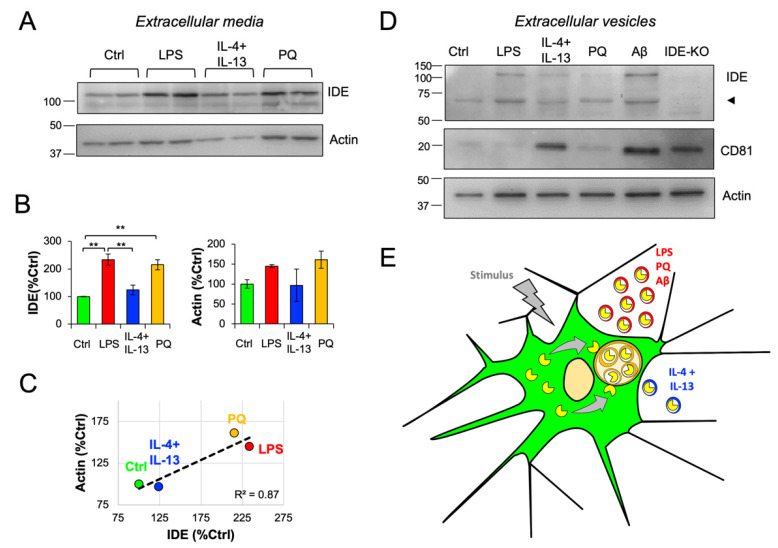
The insulin-degrading enzyme (IDE) is a zinc-dependent metalloendopeptidase that belongs to the M16A metalloprotease family. IDE is markedly expressed in the brain, where it is particularly relevant due to its in vitro amyloid beta (Aβ)-degrading activity. The subcellular localization of IDE, a paramount aspect to understand how this enzyme can perform its proteolytic functions in vivo, remains highly controversial. In this work, we addressed IDE subcellular localization from an evolutionary perspective. Phylogenetic analyses based on protein sequence and gene and protein structure were performed. An in silico analysis of IDE signal peptide suggests an evolutionary shift in IDE exportation at the prokaryote/eukaryote divide. Subcellular localization experiments in microglia revealed that IDE is mostly cytosolic. Furthermore, IDE associates to membranes by their cytoplasmatic side and further partitions between raft and non-raft domains. When stimulated, microglia change into a secretory active state, produces numerous multivesicular bodies and IDE associates with their membranes. The subsequent inward budding of such membranes internalizes IDE in intraluminal vesicles, which later allows IDE to be exported outside the cells in small extracellular vesicles. We further demonstrate that such an IDE exportation mechanism is regulated by stimuli relevant for microglia in physiological conditions and upon aging and neurodegeneration.
Keywords: amyloid β; extracellular vesicles; inflammatory state; insulin-degrading enzyme; intron-exon structure; lipid rafts; microglia; molecular evolution; oxidative stress; phylogeny.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
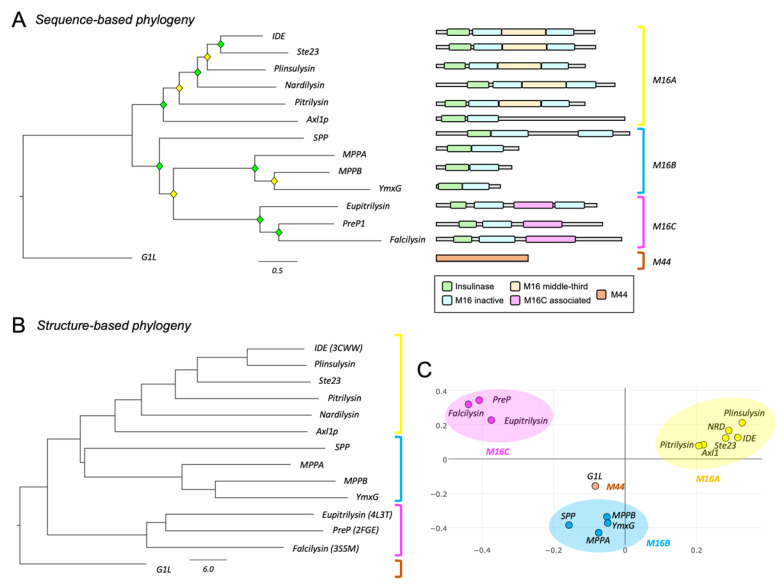
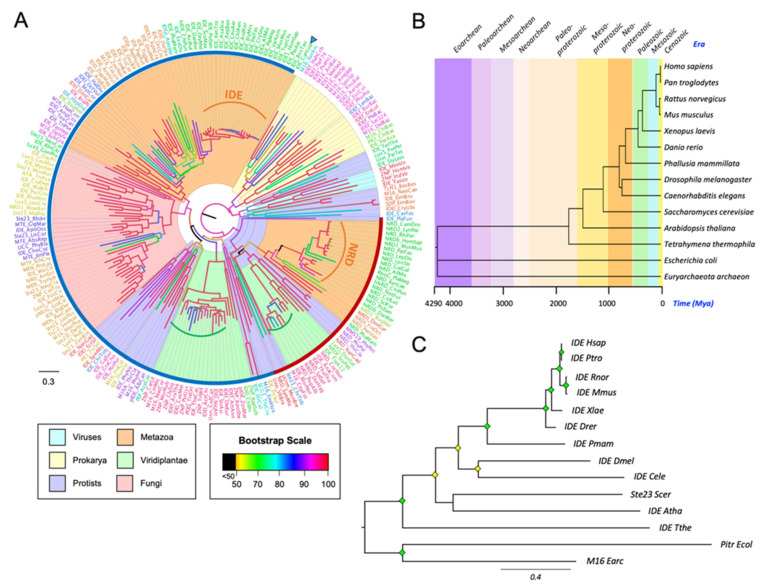
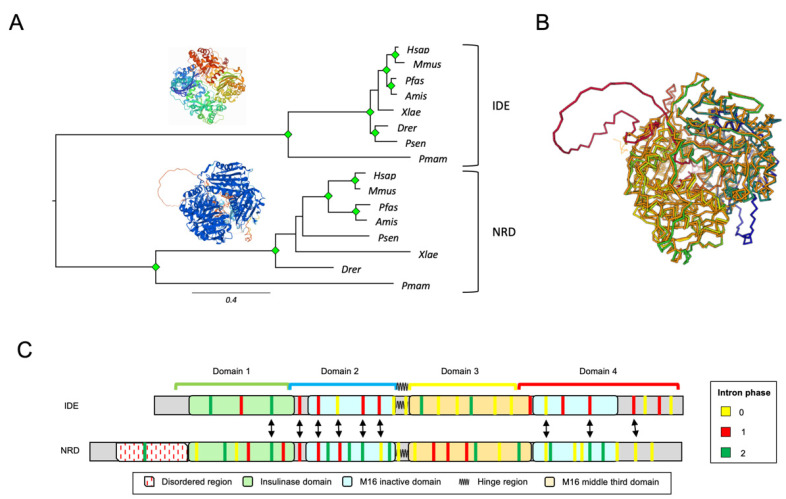
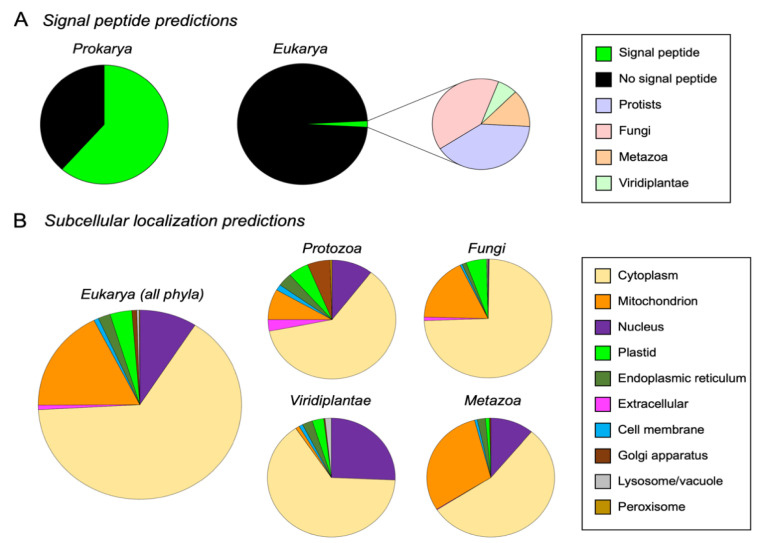
References
-
- Broh-Kahn R., Mirsky I. The Inactivation of Insulin by Tissue Extracts; the Effect of Fasting on the Insulinase Content of Rat Liver. Arch. Biochem. 1949;20:10–14. - PubMed
-
- Rawlings N.D., Barrett A.J. Handbook of Proteolytic Enzymes. Elsevier; Amsterdam, The Netherlands: 2013. Introduction: Metallopeptidases and Their Clans; pp. 325–370.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
