

Dissecting the molecular mechanisms of mitochondrial import and maturation of peroxiredoxins from yeast and mammalian cells
- PMID: 35059022
- PMCID: PMC8724339
- DOI: 10.1007/s12551-021-00899-2
Dissecting the molecular mechanisms of mitochondrial import and maturation of peroxiredoxins from yeast and mammalian cells
Abstract
Peroxiredoxins (Prxs) are cysteine-based peroxidases that play a central role in keeping the H2O2 at physiological levels. Eukaryotic cells express different Prxs isoforms, which differ in their subcellular locations and substrate specificities. Mitochondrial Prxs are synthesized in the cytosol as precursor proteins containing N-terminal cleavable presequences that act as mitochondrial targeting signals. Due to the fact that presequence controls the import of the vast majority of mitochondrial matrix proteins, the mitochondrial Prxs were initially predicted to be localized exclusively in the matrix. However, recent studies showed that mitochondrial Prxs are also targeted to the intermembrane space by mechanisms that remain poorly understood. While in yeast the IMP complex can translocate Prx1 to the intermembrane space, the maturation of yeast Prx1 and mammalian Prdx3 and Prdx5 in the matrix has been associated with sequential cleavages of the presequence by MPP and Oct1/MIP proteases. In this review, we describe the state of the art of the molecular mechanisms that control the mitochondrial import and maturation of Prxs of yeast and human cells. Once mitochondria are considered the major intracellular source of H2O2, understanding the mitochondrial Prx biogenesis pathways is essential to increase our knowledge about the H2O2-dependent cellular signaling, which is relevant to the pathophysiology of some human diseases.
Keywords: H2O2; Intermembrane space; Matrix; Mitochondria; Peroxiredoxin; Presequence.
© International Union for Pure and Applied Biophysics (IUPAB) and Springer-Verlag GmbH Germany, part of Springer Nature 2021.
Conflict of interest statement
Conflict of interestThe authors declare no competing interests.
Figures
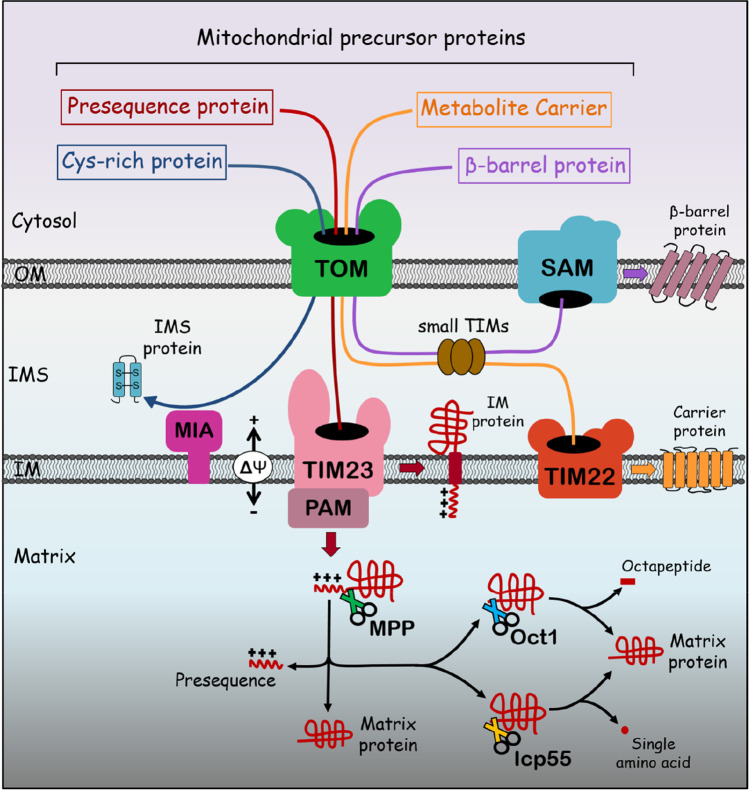

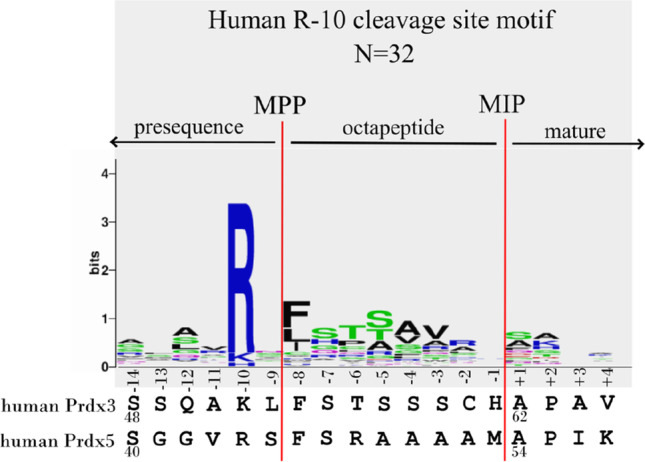
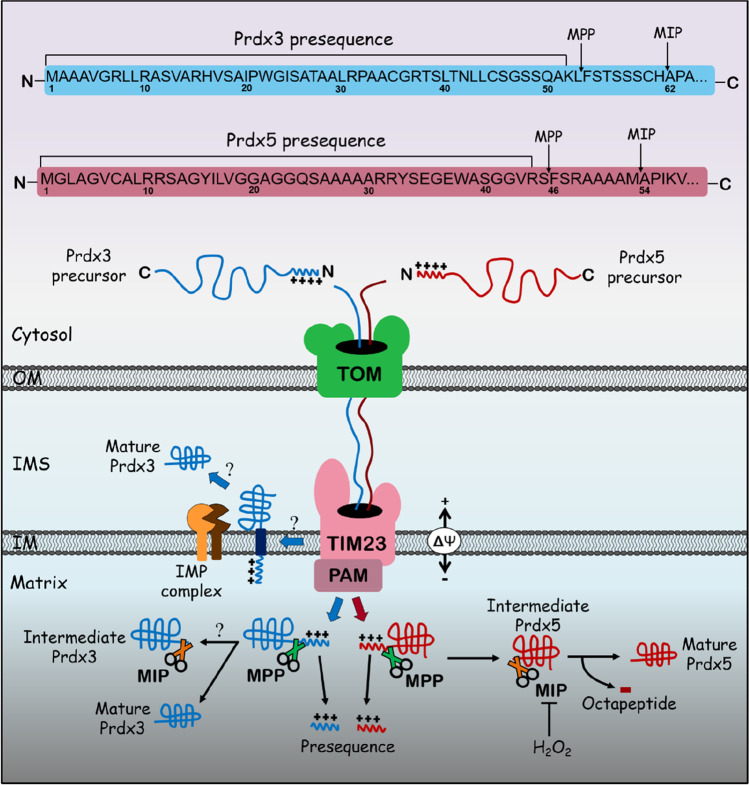
References
Publication types
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
