

Specific Human ATR and ATM Inhibitors Modulate Single Strand DNA Formation in Leishmania major Exposed to Oxidative Agent
- PMID: 35059327
- PMCID: PMC8763966
- DOI: 10.3389/fcimb.2021.802613
Specific Human ATR and ATM Inhibitors Modulate Single Strand DNA Formation in Leishmania major Exposed to Oxidative Agent
Abstract
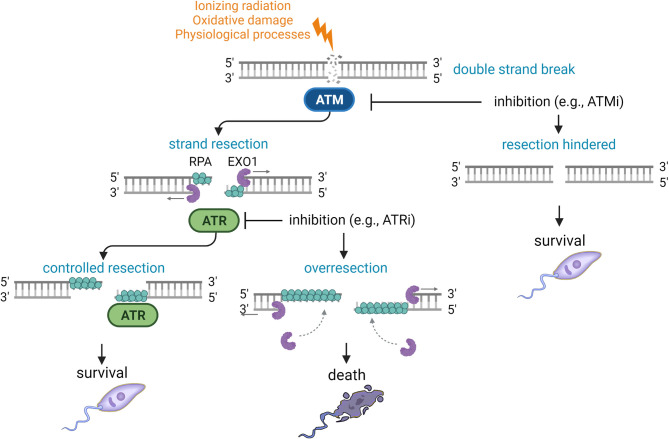
Leishmania parasites are the causative agents of a group of neglected tropical diseases known as leishmaniasis. The molecular mechanisms employed by these parasites to adapt to the adverse conditions found in their hosts are not yet completely understood. DNA repair pathways can be used by Leishmania to enable survival in the interior of macrophages, where the parasite is constantly exposed to oxygen reactive species. In higher eukaryotes, DNA repair pathways are coordinated by the central protein kinases ataxia telangiectasia mutated (ATM) and ataxia telangiectasia and Rad3 related (ATR). The enzyme Exonuclease-1 (EXO1) plays important roles in DNA replication, repair, and recombination, and it can be regulated by ATM- and ATR-mediated signaling pathways. In this study, the DNA damage response pathways in promastigote forms of L. major were investigated using bioinformatics tools, exposure of lineages to oxidizing agents and radiation damage, treatment of cells with ATM and ATR inhibitors, and flow cytometry analysis. We demonstrated high structural and important residue conservation for the catalytic activity of the putative LmjEXO1. The overexpression of putative LmjEXO1 made L. major cells more susceptible to genotoxic damage, most likely due to the nuclease activity of this enzyme and the occurrence of hyper-resection of DNA strands. These cells could be rescued by the addition of caffeine or a selective ATM inhibitor. In contrast, ATR-specific inhibition made the control cells more susceptible to oxidative damage in an LmjEXO1 overexpression-like manner. We demonstrated that ATR-specific inhibition results in the formation of extended single-stranded DNA, most likely due to EXO1 nucleasic activity. Antagonistically, ATM inhibition prevented single-strand DNA formation, which could explain the survival phenotype of lineages overexpressing LmjEXO1. These results suggest that an ATM homolog in Leishmania could act to promote end resection by putative LmjEXO1, and an ATR homologue could prevent hyper-resection, ensuring adequate repair of the parasite DNA.
Keywords: DNA repair; Leishmania major (L. major); ataxia telangiectasia and Rad3 related kinase (ATR); ataxia telangiectasia mutated (ATM); exonuclease 1.
Copyright © 2022 da Silva, Bertoldo, Naves, de Vito, Damasceno, Tosi, Machado and Pedrosa.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures


Similar articles
-
Selective human inhibitors of ATR and ATM render Leishmania major promastigotes sensitive to oxidative damage.PLoS One. 2018 Sep 28;13(9):e0205033. doi: 10.1371/journal.pone.0205033. eCollection 2018. PLoS One. 2018. PMID: 30265735 Free PMC article.
-
DNA-damage-induced degradation of EXO1 exonuclease limits DNA end resection to ensure accurate DNA repair.J Biol Chem. 2017 Jun 30;292(26):10779-10790. doi: 10.1074/jbc.M116.772475. Epub 2017 May 17. J Biol Chem. 2017. PMID: 28515316 Free PMC article.
-
Investigation of switch from ATM to ATR signaling at the sites of DNA damage induced by low and high LET radiation.DNA Repair (Amst). 2013 Dec;12(12):1143-51. doi: 10.1016/j.dnarep.2013.10.004. Epub 2013 Nov 12. DNA Repair (Amst). 2013. PMID: 24238855
-
The development of ataxia telangiectasia mutated kinase inhibitors.Mini Rev Med Chem. 2014;14(10):805-11. Mini Rev Med Chem. 2014. PMID: 25138084 Review.
-
The ATM-Chk2 and ATR-Chk1 pathways in DNA damage signaling and cancer.Adv Cancer Res. 2010;108:73-112. doi: 10.1016/B978-0-12-380888-2.00003-0. Adv Cancer Res. 2010. PMID: 21034966 Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous