

The hidden half comes into the spotlight: Peeking inside the black box of root developmental phases
- PMID: 35059627
- PMCID: PMC8760039
- DOI: 10.1016/j.xplc.2021.100246
The hidden half comes into the spotlight: Peeking inside the black box of root developmental phases
Abstract
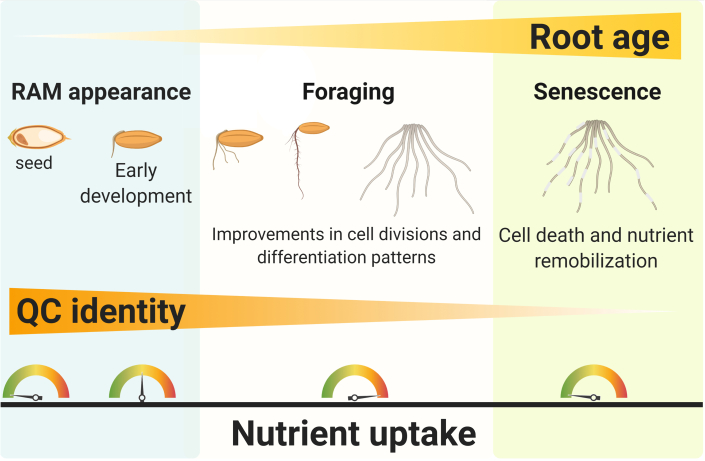
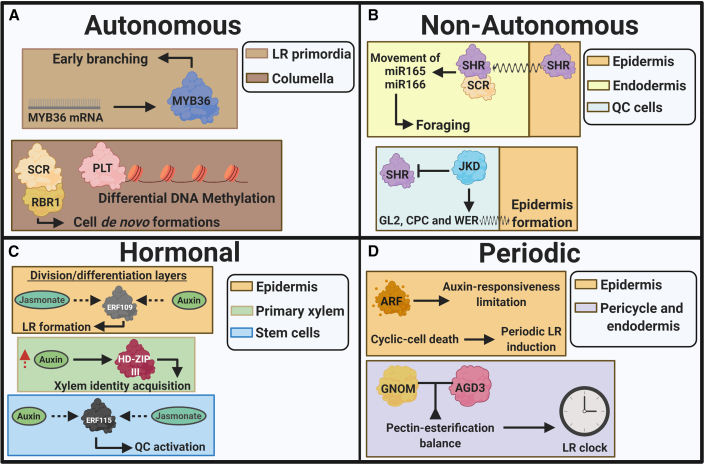
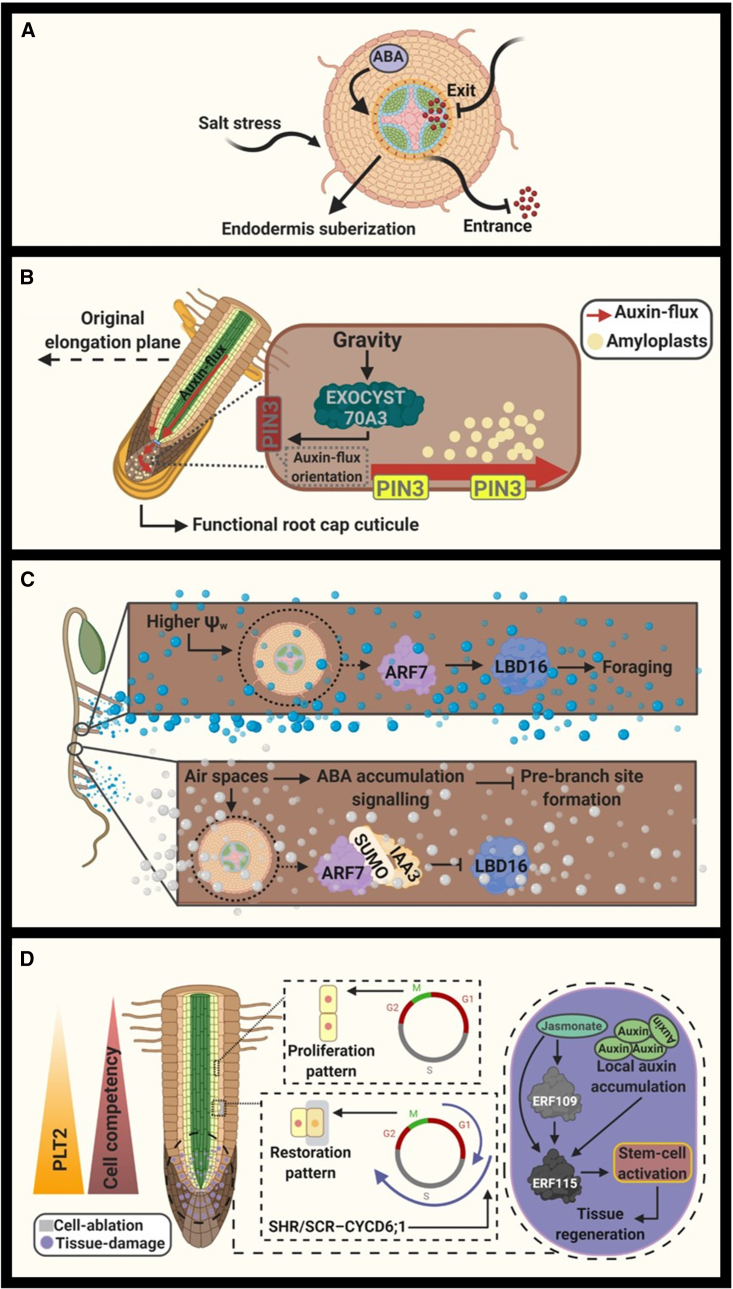
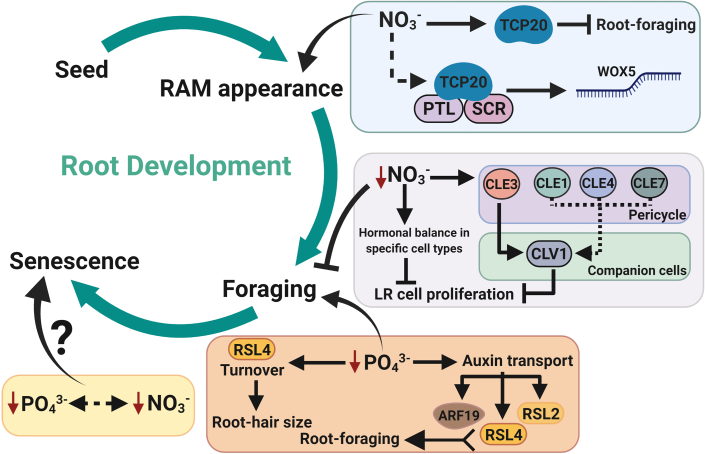
Efficient use of natural resources (e.g., light, water, and nutrients) can be improved with a tailored developmental program that maximizes the lifetime and fitness of plants. In plant shoots, a developmental phase represents a time window in which the meristem triggers the development of unique morphological and physiological traits, leading to the emergence of leaves, flowers, and fruits. Whereas developmental phases in plant shoots have been shown to enhance food production in crops, this phenomenon has remained poorly investigated in roots. In light of recent advances, we suggest that root development occurs in three main phases: root apical meristem appearance, foraging, and senescence. We provide compelling evidence suggesting that these phases are regulated by at least four developmental pathways: autonomous, non-autonomous, hormonal, and periodic. Root developmental pathways differentially coordinate organ plasticity, promoting morphological alterations, tissue regeneration, and cell death regulation. Furthermore, we suggest how nutritional checkpoints may allow progression through the developmental phases, thus completing the root life cycle. These insights highlight novel and exciting advances in root biology that may help maximize the productivity of crops through more sustainable agriculture and the reduced use of chemical fertilizers.
Keywords: cell fates; developmental transitions; nutritional checkpoints; plasticity; root clock; root development.
© 2021 The Author(s).
Figures
Similar articles
-
Lycopodium root meristem dynamics supports homology between shoots and roots in lycophytes.New Phytol. 2021 Jan;229(1):460-468. doi: 10.1111/nph.16814. Epub 2020 Aug 13. New Phytol. 2021. PMID: 32696978
-
Hormone interactions at the root apical meristem.Plant Mol Biol. 2009 Mar;69(4):383-96. doi: 10.1007/s11103-008-9393-6. Epub 2008 Sep 19. Plant Mol Biol. 2009. PMID: 18807199 Review.
-
Developmental anatomy and branching of roots of four Zeylanidium species (podostemaceae), with implications for evolution of foliose roots.Ann Bot. 2002 Dec;90(6):735-44. doi: 10.1093/aob/mcf259. Ann Bot. 2002. PMID: 12451029 Free PMC article.
-
Developmental anatomy of seedlings of Indodalzellia gracilis (Podostemaceae).Plant Biol (Stuttg). 2010 Sep 1;12(5):794-9. doi: 10.1111/j.1438-8677.2009.00275.x. Plant Biol (Stuttg). 2010. PMID: 20701703
-
Redox regulation of root apical meristem organization: connecting root development to its environment.Plant Physiol Biochem. 2010 May;48(5):328-36. doi: 10.1016/j.plaphy.2009.11.005. Epub 2009 Dec 3. Plant Physiol Biochem. 2010. PMID: 20031434 Review.
Cited by
-
Genome-wide identification of bZIP transcription factors and their expression analysis in Platycodon grandiflorus under abiotic stress.Front Plant Sci. 2024 May 28;15:1403220. doi: 10.3389/fpls.2024.1403220. eCollection 2024. Front Plant Sci. 2024. PMID: 38863542 Free PMC article.
-
Xylooligosaccharides Enhance Lettuce Root Morphogenesis and Growth Dynamics.Plants (Basel). 2024 Jun 19;13(12):1699. doi: 10.3390/plants13121699. Plants (Basel). 2024. PMID: 38931130 Free PMC article.
-
The MADS-box gene XAANTAL1 participates in Arabidopsis thaliana primary root growth and columella stem cell patterns in response to ROS, via direct regulation of PEROXIDASE 28 and RETINOBLASTOMA-RELATED genes.J Exp Bot. 2025 Jan 10;76(2):411-432. doi: 10.1093/jxb/erae415. J Exp Bot. 2025. PMID: 39377268 Free PMC article.
-
SPL13 controls a root apical meristem phase change by triggering oriented cell divisions.Science. 2024 Nov 15;386(6723):eado4298. doi: 10.1126/science.ado4298. Epub 2024 Nov 15. Science. 2024. PMID: 39541454 Free PMC article.
References
-
- Araya T., Miyamoto M., Wibowo J., Suzuki A., Kojima S., Tsuchiya Y.N., Sawa S., Fukuda H., von Wirén N., Takahashi H. CLE-CLAVATA1 peptide-receptor signaling module regulates the expansion of plant root systems in a nitrogen-dependent manner. Proc. Natl. Acad. Sci. U S A. 2014;111:2029–2034. - PMC - PubMed
-
- Bagniewska-Zadworna A., Arasimowicz-Jelonek M. The mystery of underground death: cell death in roots during ontogeny and in response to environmental factors. Plant Biol. 2016;18:171–184. - PubMed
-
- Bagniewska-Zadworna A., Stelmasik A., Minicka J. From birth to death—Populus trichocarpa fibrous roots functional anatomy. Biol. Plantarum. 2014;58:551–560.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
