

Nampt activator P7C3 ameliorates diabetes and improves skeletal muscle function modulating cell metabolism and lipid mediators
- PMID: 35060352
- PMCID: PMC8977983
- DOI: 10.1002/jcsm.12887
Nampt activator P7C3 ameliorates diabetes and improves skeletal muscle function modulating cell metabolism and lipid mediators
Abstract
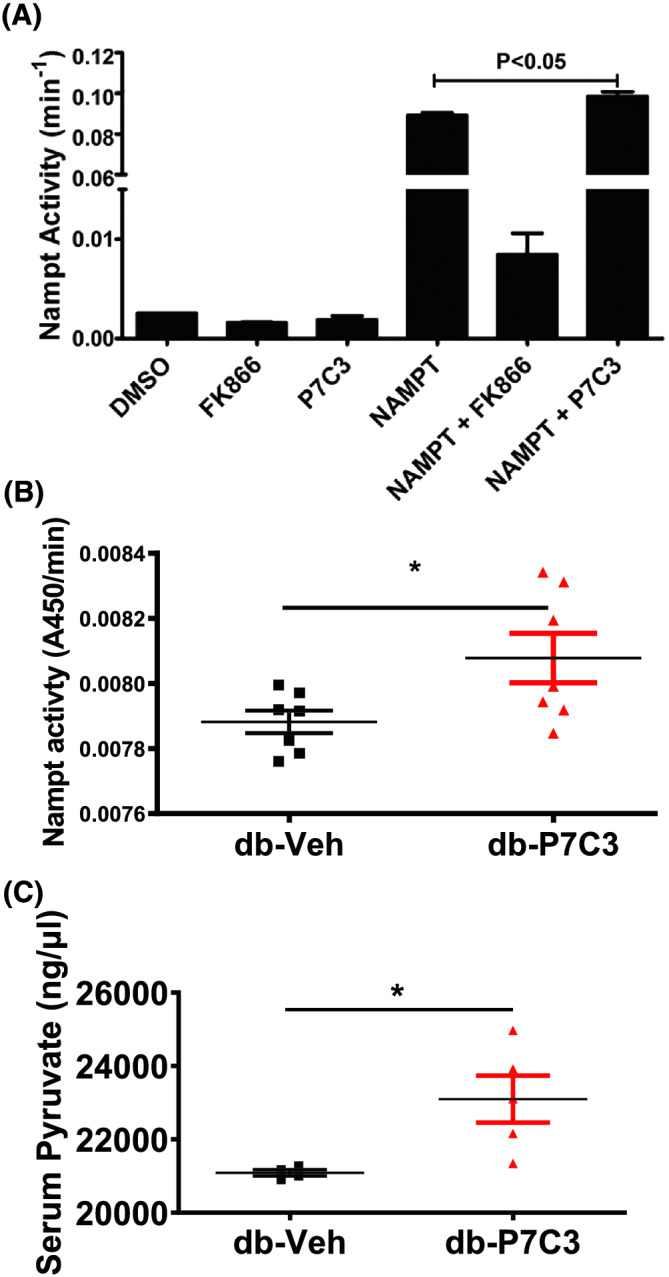
Background: Nicotinamide phosphoribosyltransferase (Nampt), a key enzyme in NAD salvage pathway is decreased in metabolic diseases, and its precise role in skeletal muscle function is not known. We tested the hypothesis, Nampt activation by P7C3 (3,6-dibromo-α-[(phenylamino)methyl]-9H-carbazol-9-ethanol) ameliorates diabetes and muscle function.
Methods: We assessed the functional, morphometric, biochemical, and molecular effects of P7C3 treatment in skeletal muscle of type 2 diabetic (db/db) mice. Nampt+/- mice were utilized to test the specificity of P7C3.
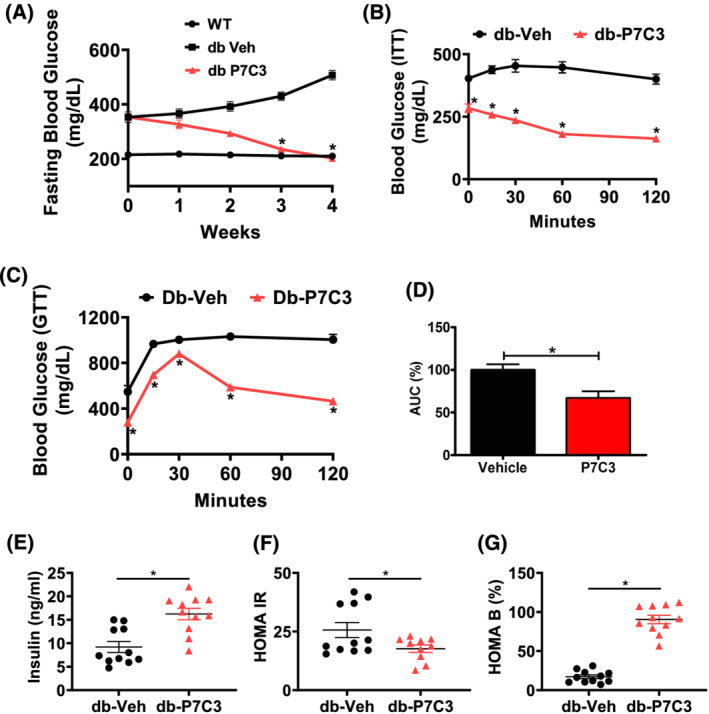
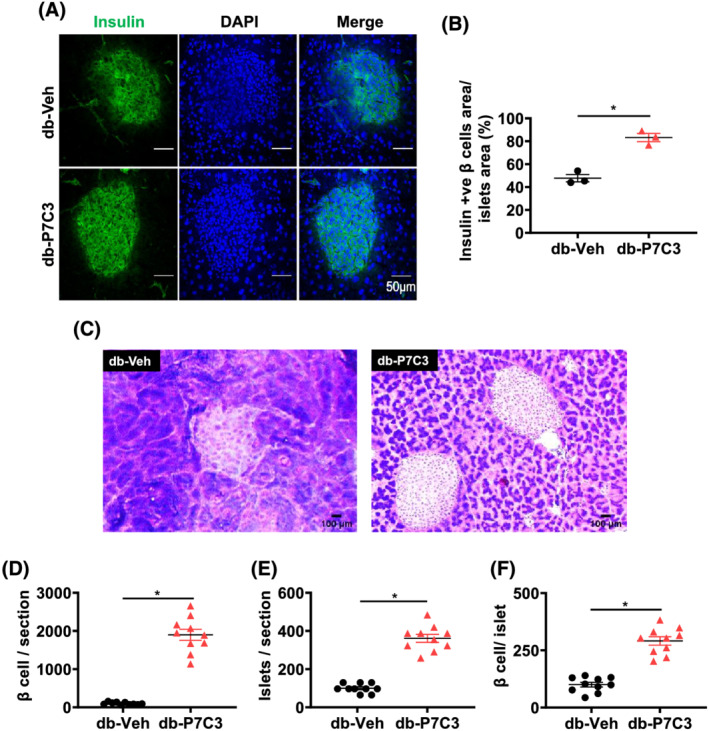
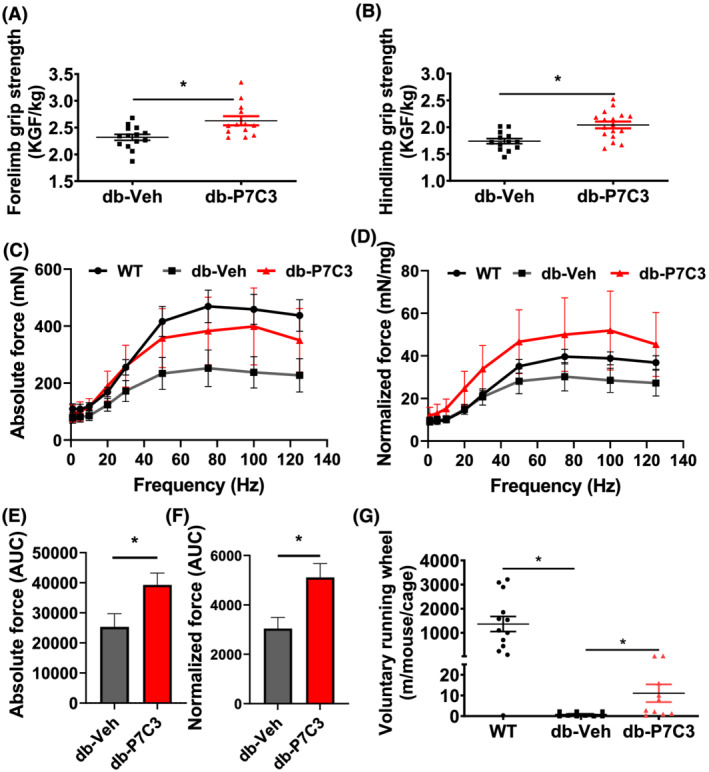
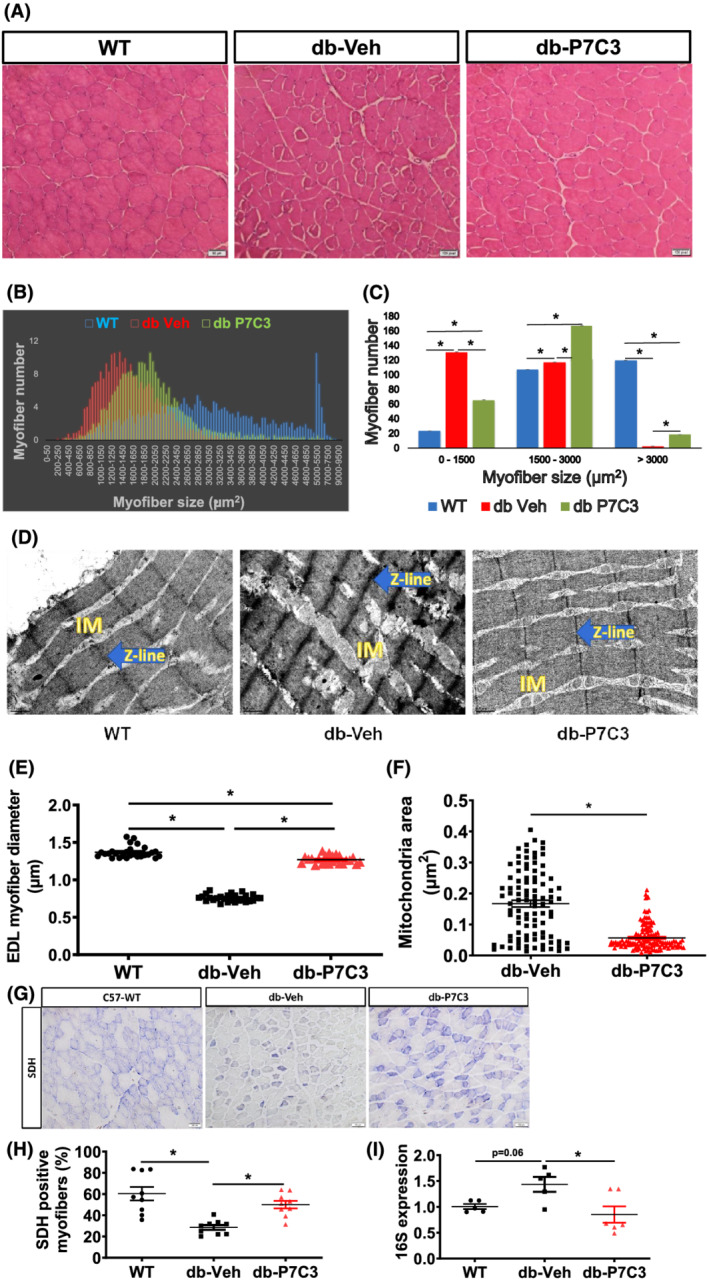
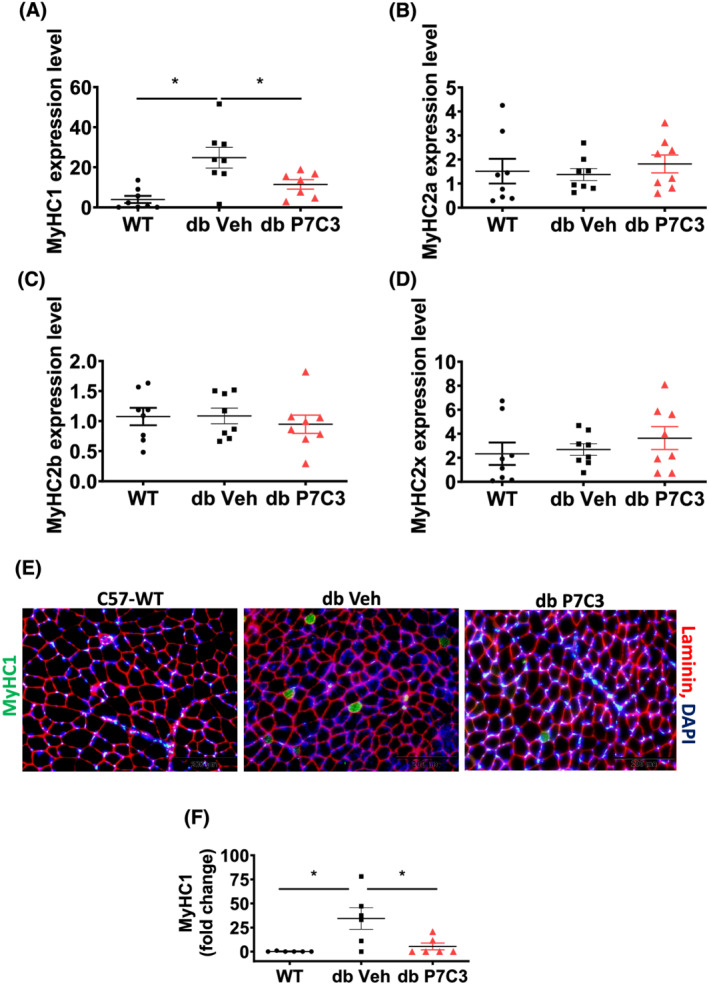
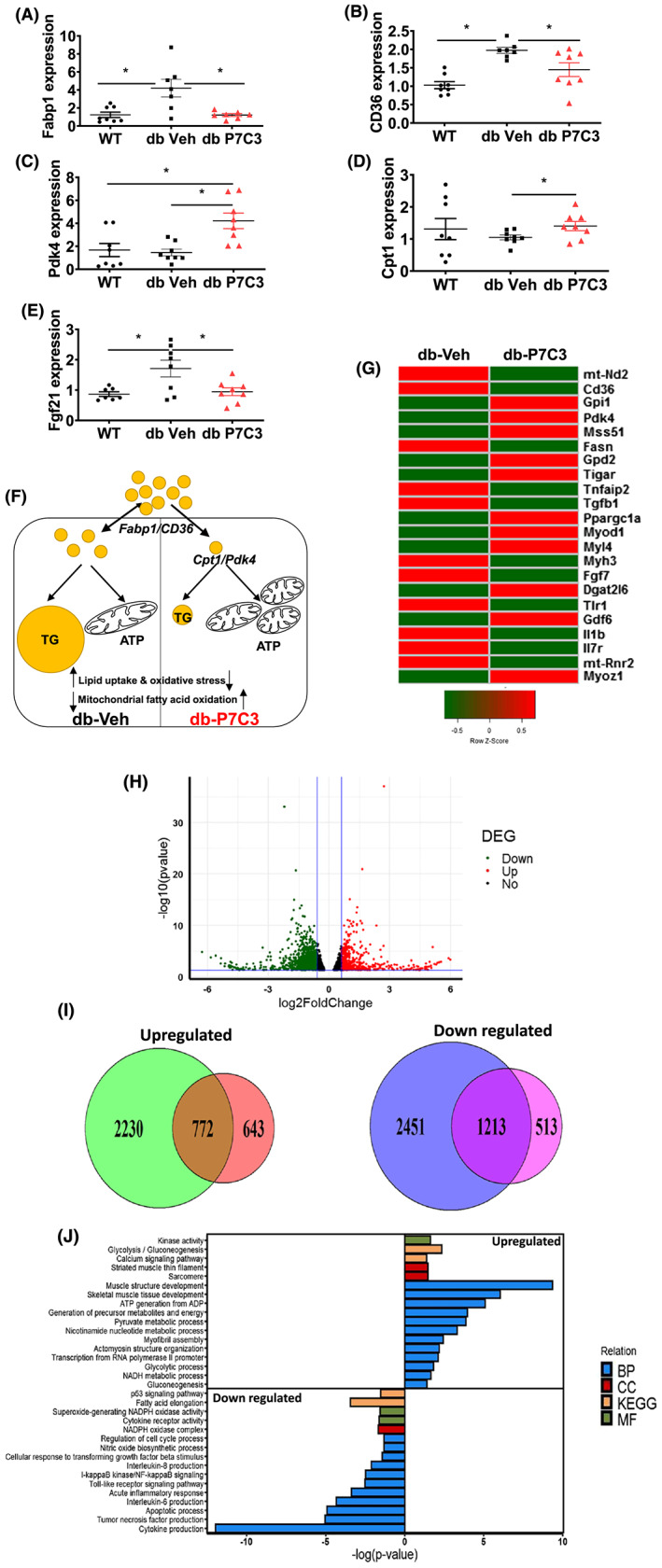
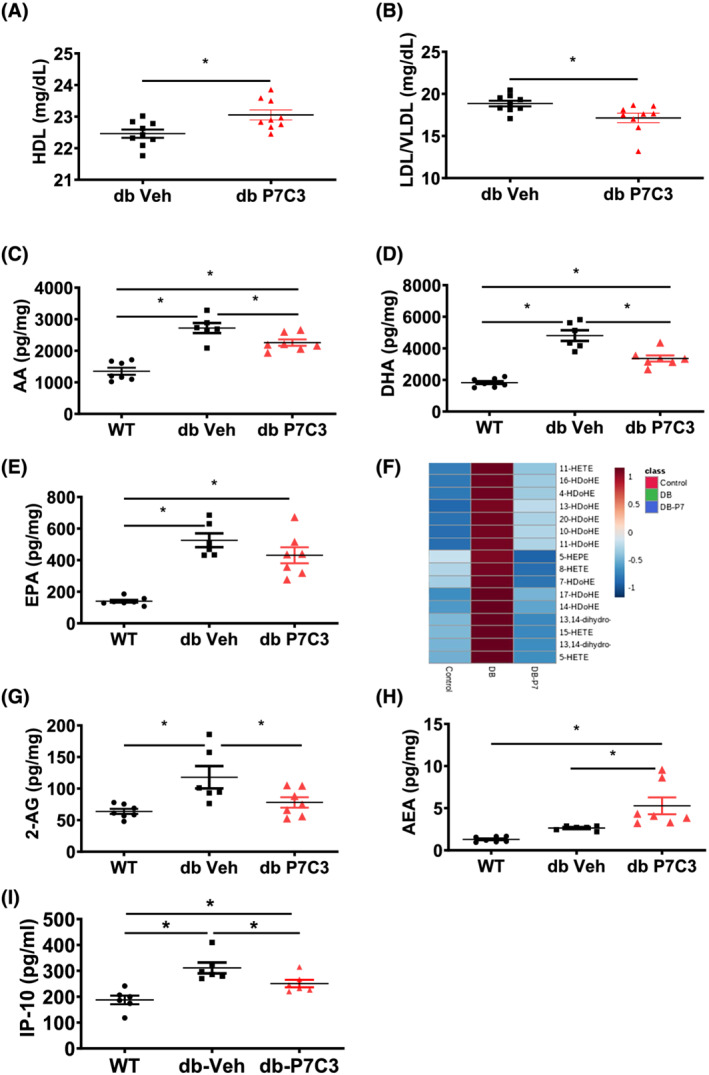
Results: Insulin resistance increased 1.6-fold in diabetic mice compared with wild-type mice and after 4 weeks treatment with P7C3 rescued diabetes (P < 0.05). In the db-P7C3 mice fasting blood glucose levels decreased to 0.96-fold compared with C57Bl/6J wild-type naïve control mice. The insulin and glucose tolerance tests blood glucose levels were decreased to 0.6-fold and 0.54-folds, respectively, at 120 min along with an increase in insulin secretion (1.76-fold) and pancreatic β-cells (3.92-fold) in db-P7C3 mice. The fore-limb and hind-limb grip strengths were increased to 1.13-fold and 1.17-fold, respectively, together with a 14.2-fold increase in voluntary running wheel distance in db-P7C3 mice. P7C3 treatment resulted in a 1.4-fold and 7.1-fold increase in medium-sized and larger-sized myofibres cross-sectional area, with a concomitant 0.5-fold decrease in smaller-sized myofibres of tibialis anterior (TA) muscle. The transmission electron microscopy images also displayed a 1.67-fold increase in myofibre diameter of extensor digitorum longus muscle along with 2.9-fold decrease in mitochondrial area in db-P7C3 mice compared with db-Veh mice. The number of SDH positive myofibres were increased to 1.74-fold in db-P7C3 TA muscles. The gastrocnemius and TA muscles displayed a decrease in slow oxidative myosin heavy chain type1 (MyHC1) myofibres expression (0.46-fold) and immunostaining (6.4-fold), respectively. qPCR analysis displayed a 2.9-fold and 1.3-fold increase in Pdk4 and Cpt1, and 0.55-fold and 0.59-fold decrease in Fgf21 and 16S in db-P7C3 mice. There was also a 3.3-fold and 1.9-fold increase in Fabp1 and CD36 in db-Veh mice. RNA-seq differential gene expression volcano plot displayed 1415 genes to be up-regulated and 1726 genes down-regulated (P < 0.05) in db-P7C3 mice. There was 1.02-fold increase in serum HDL, and 0.9-fold decrease in low-density lipoprotein/very low-density lipoprotein ratio in db-P7C3 mice. Lipid profiling of gastrocnemius muscle displayed a decrease in inflammatory lipid mediators n-6; AA (0.83-fold), and n-3; DHA (0.69-fold) and EPA (0.81-fold), and a 0.66-fold decrease in endocannabinoid 2-AG and 2.0-fold increase in AEA in db-P7C3 mice.
Conclusions: Overall, we demonstrate that P7C3 activates Nampt, improves type 2 diabetes and skeletal muscle function in db/db mice.
Keywords: Insulin sensitivity; Nampt; P7C3; Pathophysiology; Physical performance; Skeletal muscle.
© 2022 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Patent awarded to USF: US Patent 11,007,178, 2021. USF Provisional patent filed: 19A039PR.
Figures
References
-
- Wojciechowska J, Krajewski W, Bolanowski M, Kręcicki T, Zatoński T. Diabetes and cancer: a review of current knowledge. Exp Clin Endocrinol Diabetes 2016;124:263–275. - PubMed
-
- Phielix E, Mensink M. Type 2 diabetes mellitus and skeletal muscle metabolic function. Physiol Behav 2008;94:252–258. - PubMed
-
- Twig G, Zucker I, Afek A, Cukierman‐Yaffe T, Bendor CD, Derazne E, et al. Adolescent obesity and early‐onset type 2 diabetes. Diabetes Care 2020;43:1487–1495. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
