

The blood-brain barrier is dysregulated in COVID-19 and serves as a CNS entry route for SARS-CoV-2
- PMID: 35063125
- PMCID: PMC8772030
- DOI: 10.1016/j.stemcr.2021.12.011
The blood-brain barrier is dysregulated in COVID-19 and serves as a CNS entry route for SARS-CoV-2
Abstract
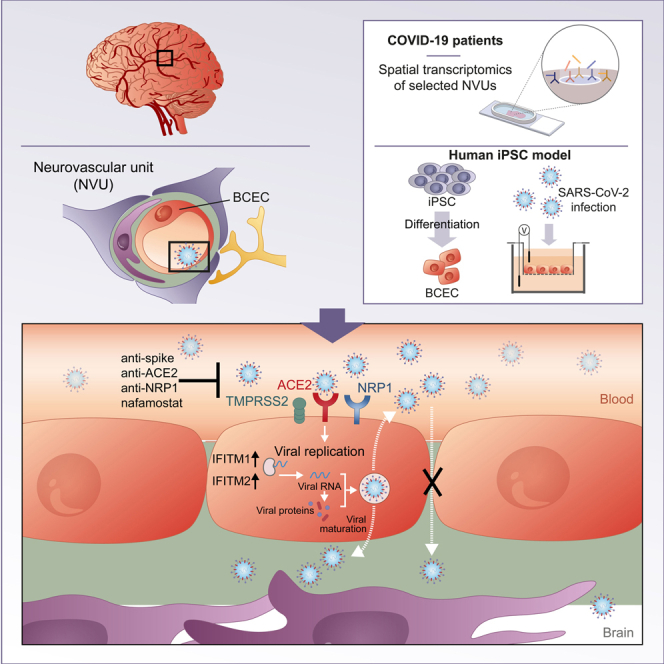
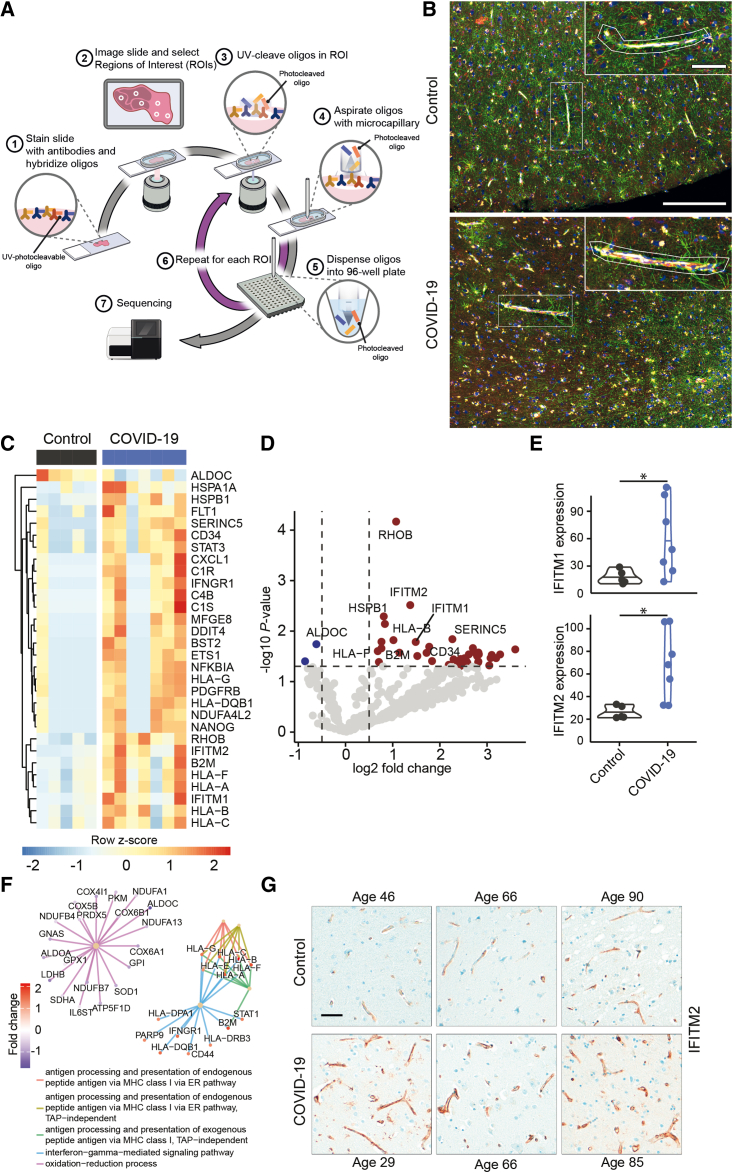
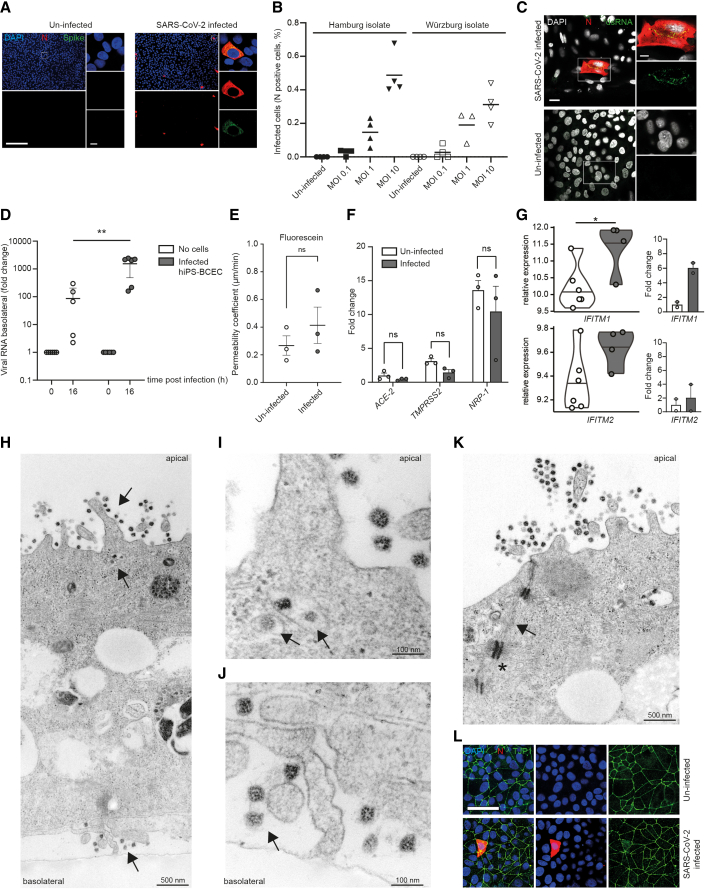
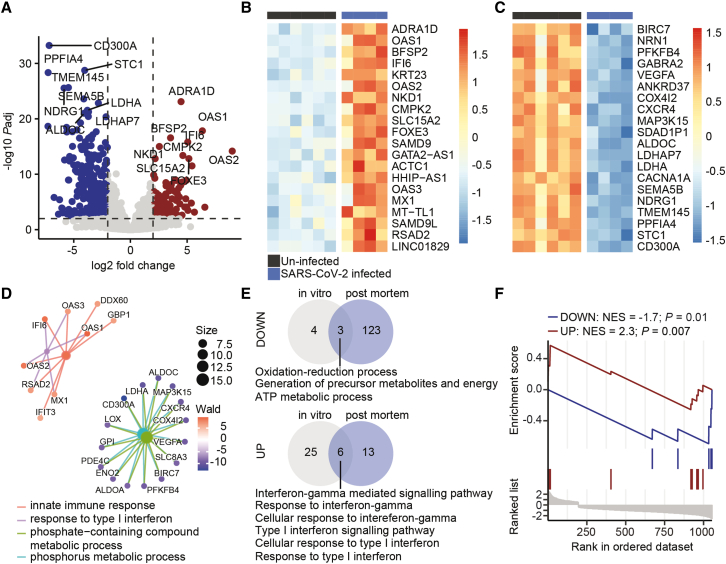
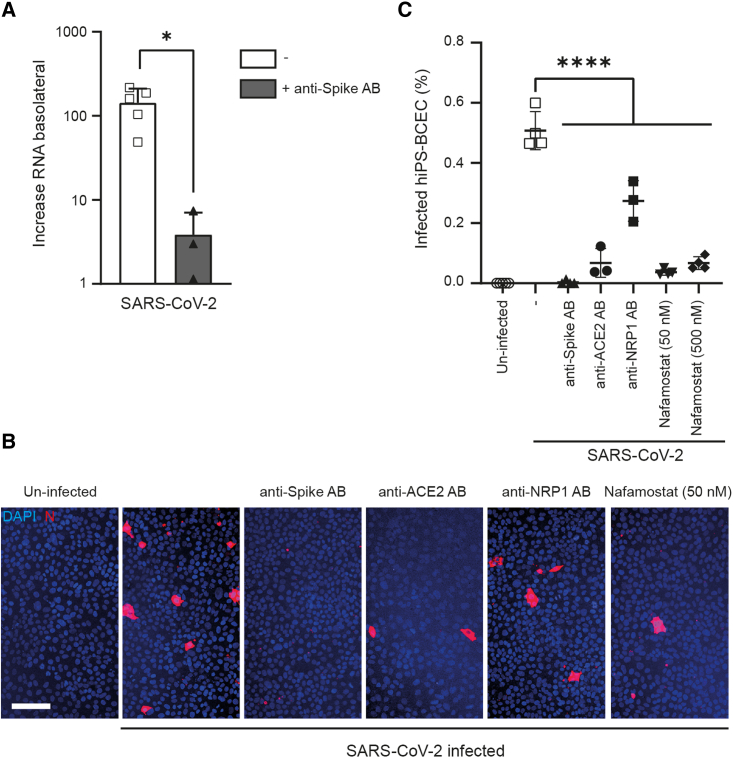
Neurological complications are common in COVID-19. Although SARS-CoV-2 has been detected in patients' brain tissues, its entry routes and resulting consequences are not well understood. Here, we show a pronounced upregulation of interferon signaling pathways of the neurovascular unit in fatal COVID-19. By investigating the susceptibility of human induced pluripotent stem cell (hiPSC)-derived brain capillary endothelial-like cells (BCECs) to SARS-CoV-2 infection, we found that BCECs were infected and recapitulated transcriptional changes detected in vivo. While BCECs were not compromised in their paracellular tightness, we found SARS-CoV-2 in the basolateral compartment in transwell assays after apical infection, suggesting active replication and transcellular transport of virus across the blood-brain barrier (BBB) in vitro. Moreover, entry of SARS-CoV-2 into BCECs could be reduced by anti-spike-, anti-angiotensin-converting enzyme 2 (ACE2)-, and anti-neuropilin-1 (NRP1)-specific antibodies or the transmembrane protease serine subtype 2 (TMPRSS2) inhibitor nafamostat. Together, our data provide strong support for SARS-CoV-2 brain entry across the BBB resulting in increased interferon signaling.
Keywords: COVID-19; SARS-CoV-2; blood-brain barrier; hiPSC; infection model; neurovascular unit.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
SARS-CoV-2 and type I interferon signaling in brain endothelial cells: Blurring the lines between friend or foe.Stem Cell Reports. 2022 May 10;17(5):1012-1013. doi: 10.1016/j.stemcr.2022.04.011. Stem Cell Reports. 2022. PMID: 35545022 Free PMC article. No abstract available.
-
Response to: SARS-CoV-2 and type I interferon signaling in brain endothelial cells: Blurring the lines between friend or foe.Stem Cell Reports. 2022 May 10;17(5):1014-1015. doi: 10.1016/j.stemcr.2022.04.012. Stem Cell Reports. 2022. PMID: 35545023 Free PMC article. No abstract available.
References
-
- Appelt-Menzel A., Cubukova A., Gunther K., Edenhofer F., Piontek J., Krause G., Stuber T., Walles H., Neuhaus W., Metzger M. Establishment of a human blood-brain barrier co-culture model mimicking the neurovascular unit using induced pluri- and multipotent stem cells. Stem Cell Rep. 2017;8:894–906. - PMC - PubMed
-
- Bao L., Deng W., Huang B., Gao H., Liu J., Ren L., Wei Q., Yu P., Xu Y., Qi F., et al. The pathogenicity of SARS-CoV-2 in hACE2 transgenic mice. Nature. 2020;583:830–833. - PubMed
-
- Buzhdygan T.P., DeOre B.J., Baldwin-Leclair A., Bullock T.A., McGary H.M., Khan J.A., Razmpour R., Hale J.F., Galie P.A., Potula R., et al. The SARS-CoV-2 spike protein alters barrier function in 2D static and 3D microfluidic in-vitro models of the human blood-brain barrier. Neurobiol. Dis. 2020;146:105131. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
