

Mechanisms and Consequences of Cerebellar Purkinje Cell Disinhibition in a Mouse Model of Duchenne Muscular Dystrophy
- PMID: 35064002
- PMCID: PMC8916753
- DOI: 10.1523/JNEUROSCI.1256-21.2022
Mechanisms and Consequences of Cerebellar Purkinje Cell Disinhibition in a Mouse Model of Duchenne Muscular Dystrophy
Abstract
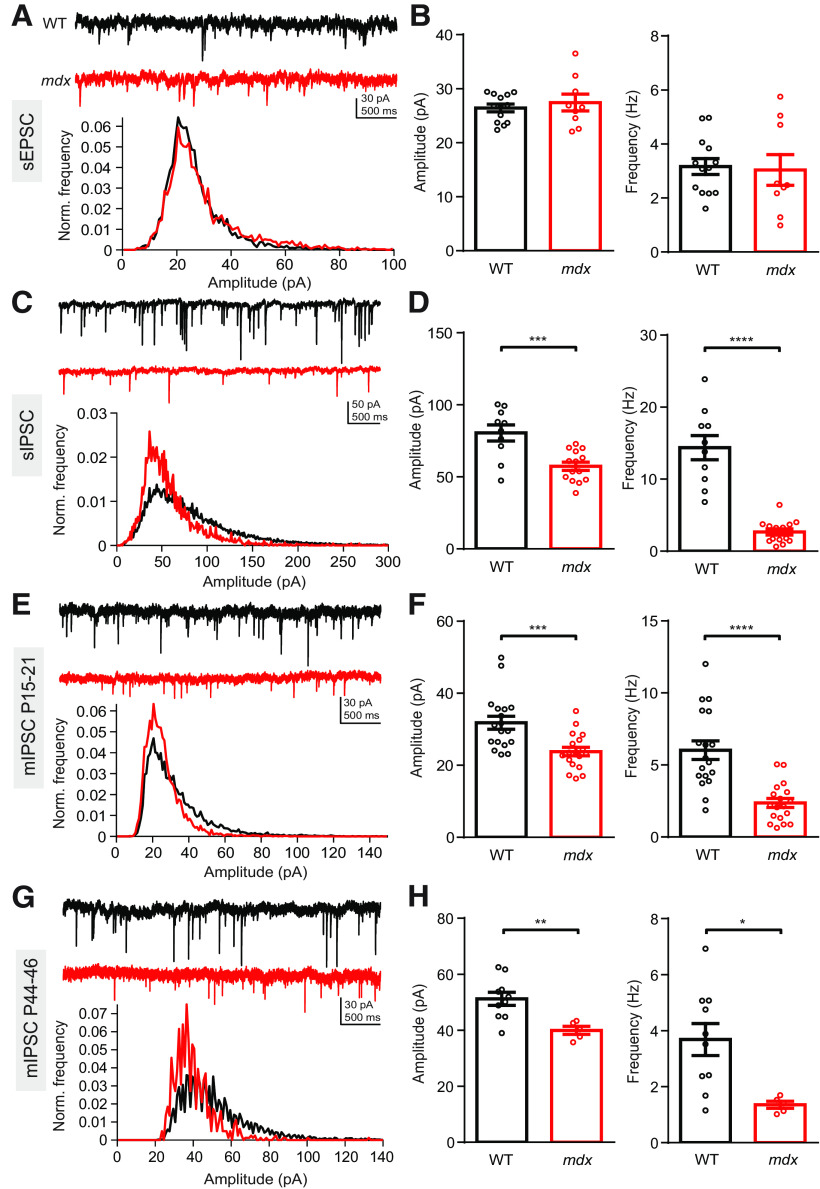
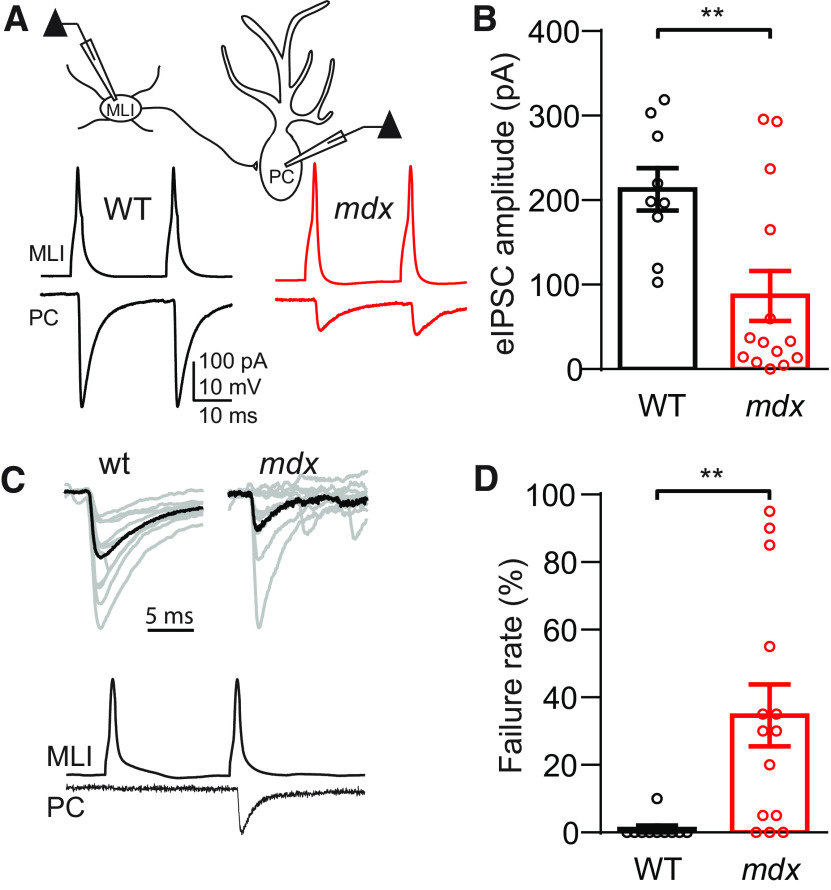
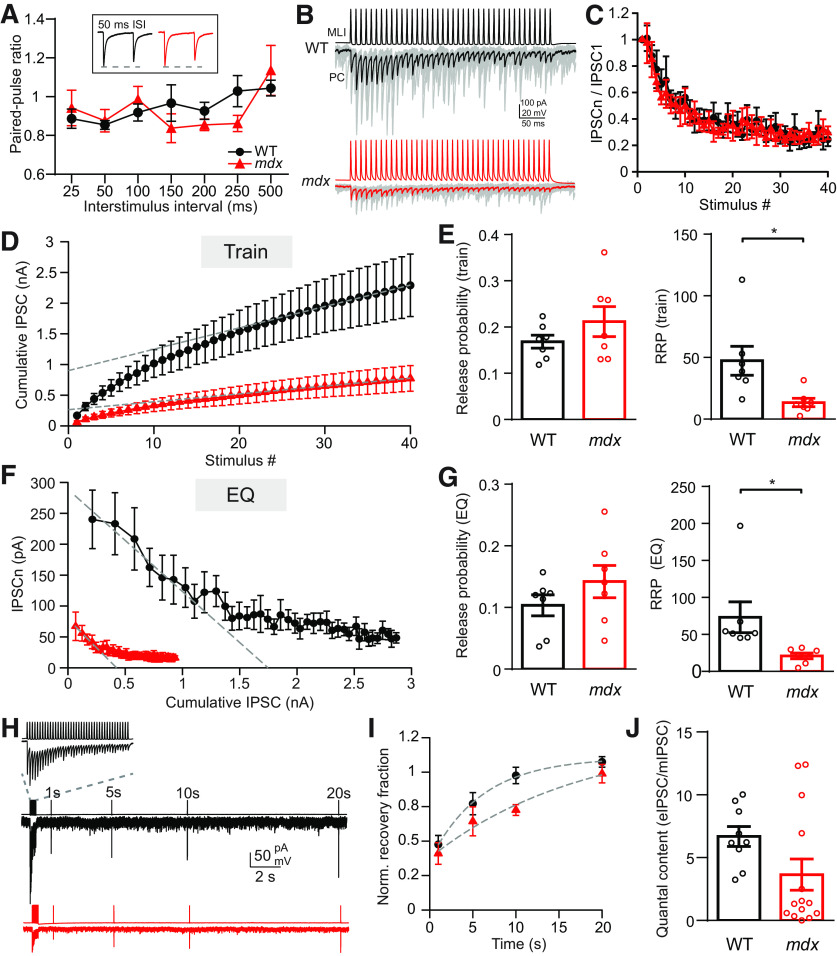
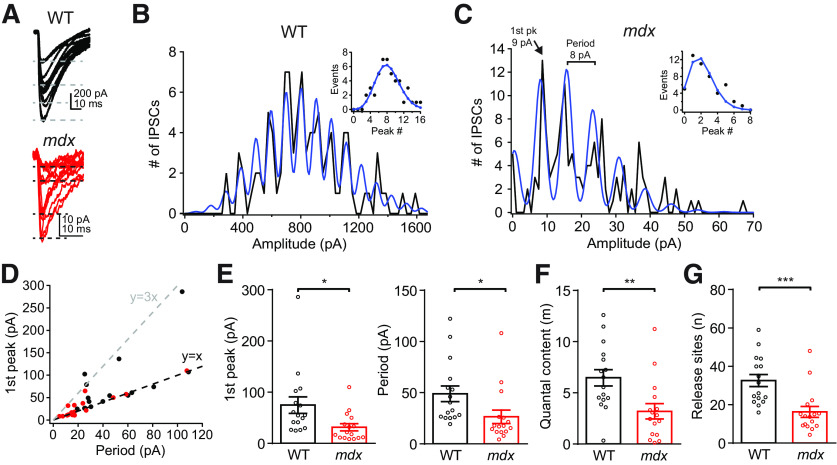
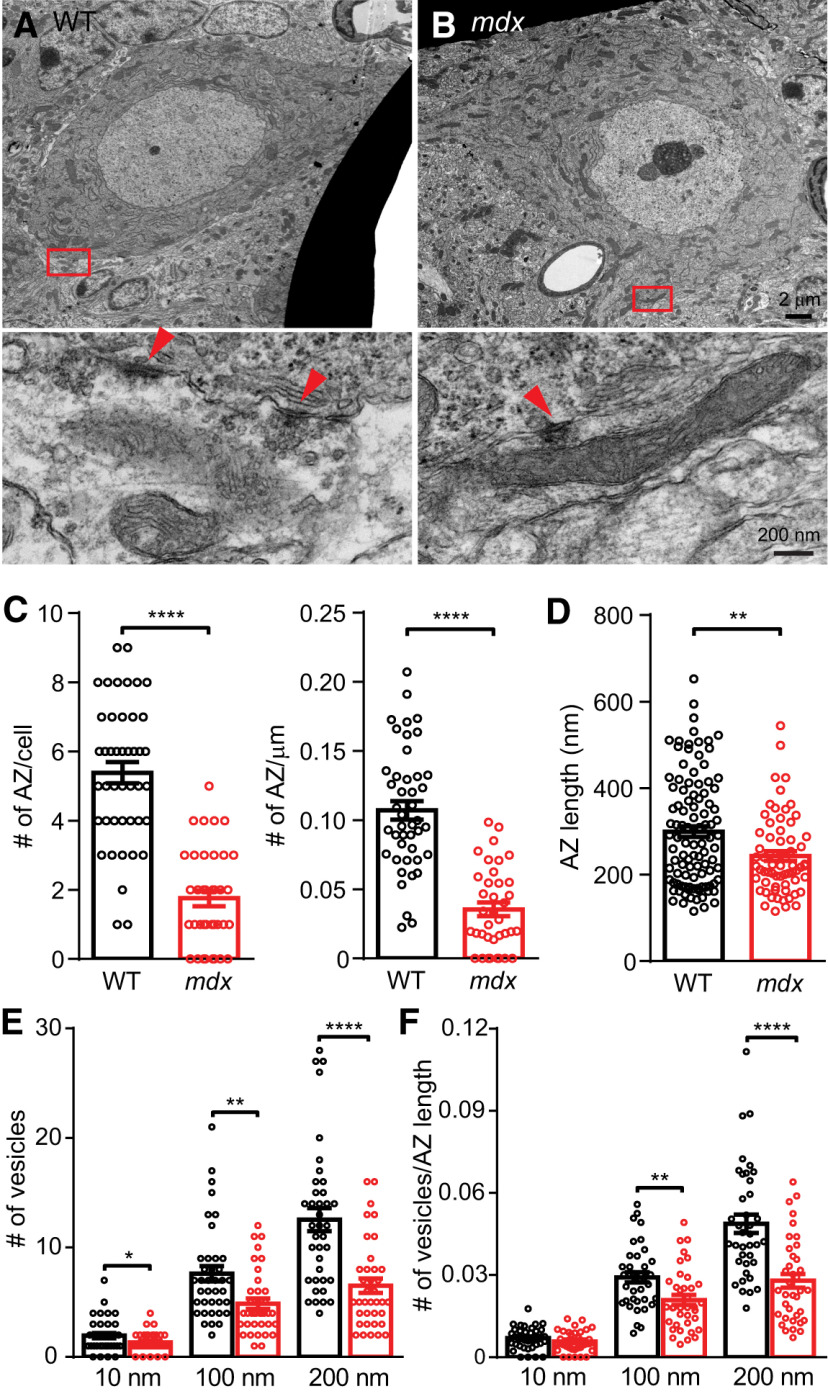
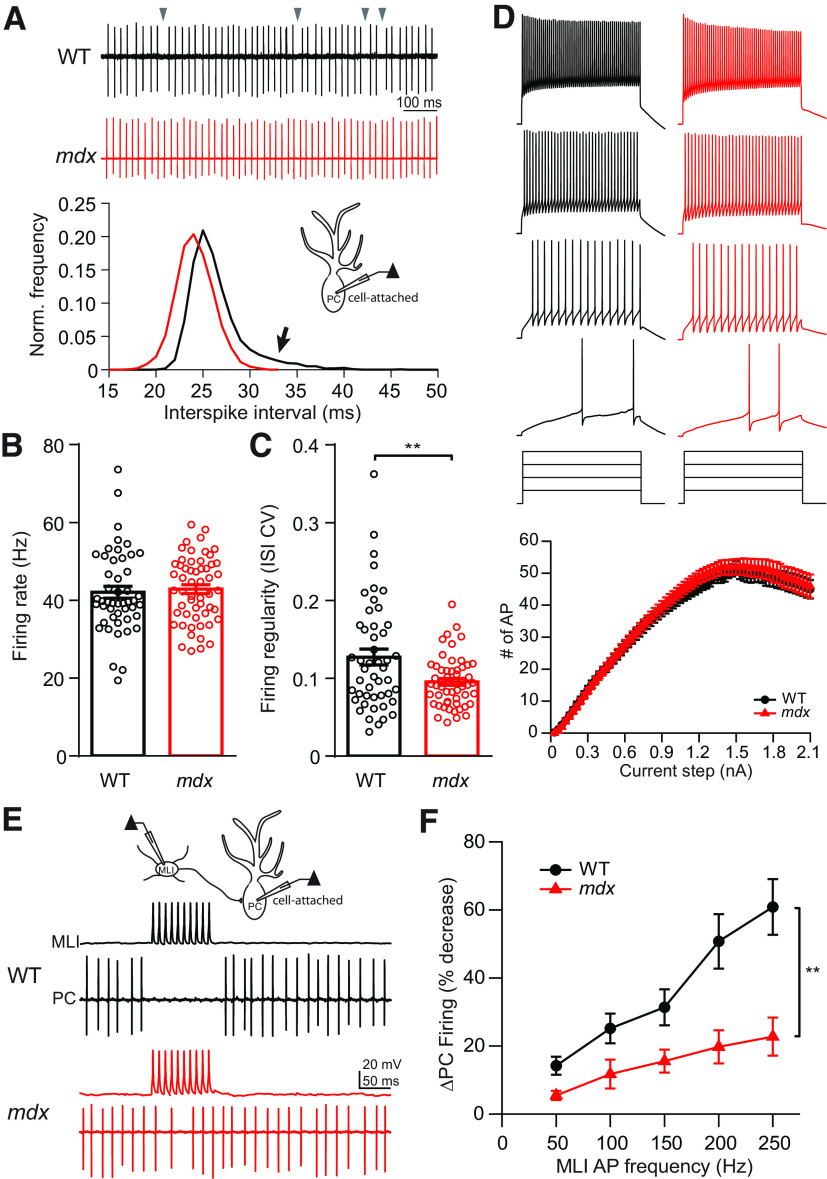
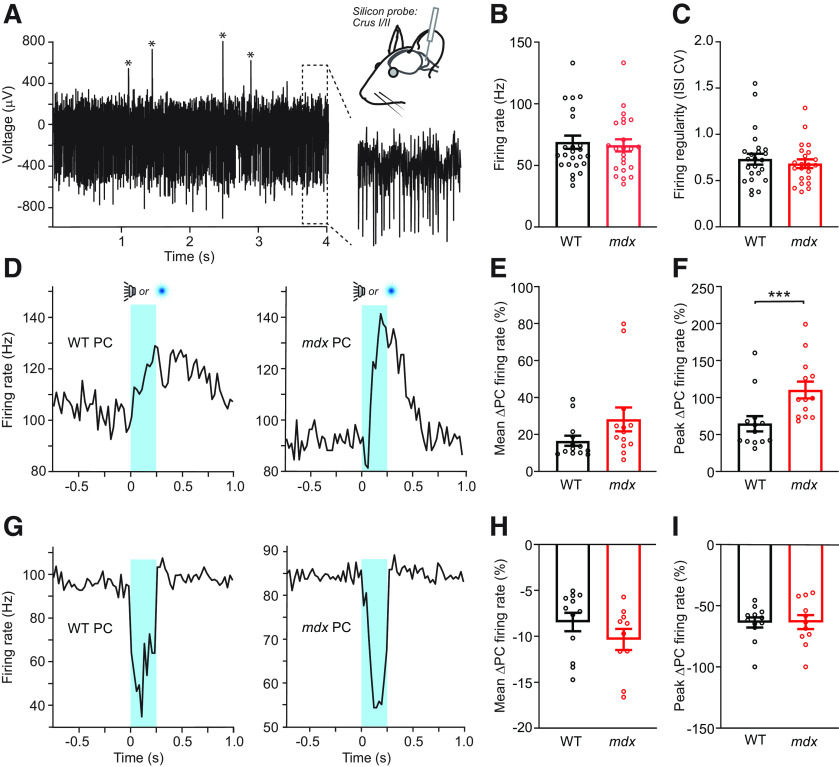
Duchenne muscular dystrophy (DMD), the most common form of childhood muscular dystrophy, is caused by mutations in the dystrophin gene. In addition to debilitating muscle degeneration, patients display a range of cognitive deficits thought to result from the loss of dystrophin normally expressed in the brain. While the function of dystrophin in muscle tissue is well characterized, its role in the brain is still poorly understood. The highest expression of dystrophin in the mouse brain is in cerebellar Purkinje cells (PCs), where it colocalizes with GABAA receptor clusters. Using ex vivo electrophysiological recordings from connected molecular layer interneuron (MLI)-PC pairs, we investigated changes in inhibitory synaptic transmission caused by dystrophin deficiency. In male mdx mice (which lack long-form dystrophin), we found that responses at MLI-PC pairs were reduced by ∼60% because of both decreased quantal response amplitude and a reduced number of functional vesicle release sites. Using electron microscopy, we found significantly fewer and smaller anatomically defined inhibitory synapses contacting the soma of PCs in mdx mice, suggesting that dystrophin may play a critical role in synapse formation and/or maintenance. Functionally, we found reduced MLI-evoked pauses in PC firing in acute slices. In vivo recordings from awake mdx mice showed increased sensory-evoked simple spike firing in positively modulating PCs, consistent with reduced feedforward inhibition, but no change in negatively modulating PCs. These data suggest that dystrophin deficiency in PCs disrupts inhibitory signaling in the cerebellar circuit and PC firing patterns, potentially contributing to cognitive and motor deficits observed in mdx mice and DMD patients.SIGNIFICANCE STATEMENT Duchenne muscular dystrophy (DMD) is primarily characterized by progressive muscle weakening caused by genetic mutations in the gene for dystrophin. Dystrophin is also normally expressed in the CNS, and DMD patients experience a range of nonprogressive cognitive deficits. The pathophysiology of CNS neurons resulting from loss of dystrophin and the function of dystrophin in neurons are still poorly understood. Using cerebellar PCs as a model, we found that the loss of dystrophin specifically disrupts the number and strength of inhibitory synaptic connections, suggesting that dystrophin participates in formation and/or maintenance of these synapses. This work provides insight into the function of dystrophin in the CNS and establishes neuronal and synaptic dysfunction, which may underlie cognitive deficits in DMD.
Keywords: DMD; Purkinje; dystrophin; inhibition; mdx; synapse.
Copyright © 2022 the authors.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases