

The microbiota regulates hematopoietic stem cell fate decisions by controlling iron availability in bone marrow
- PMID: 35065706
- PMCID: PMC8818037
- DOI: 10.1016/j.stem.2021.12.009
The microbiota regulates hematopoietic stem cell fate decisions by controlling iron availability in bone marrow
Abstract
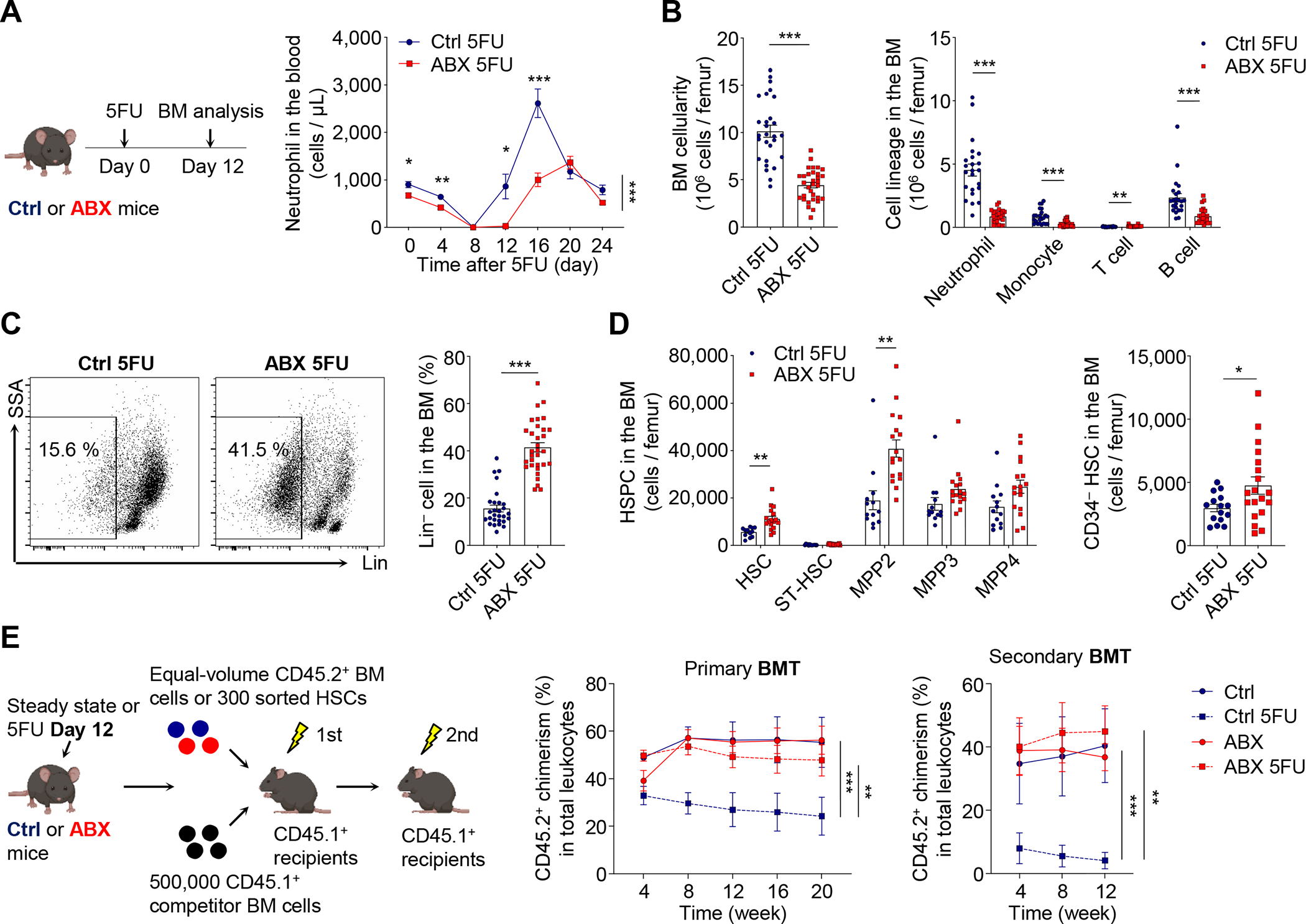
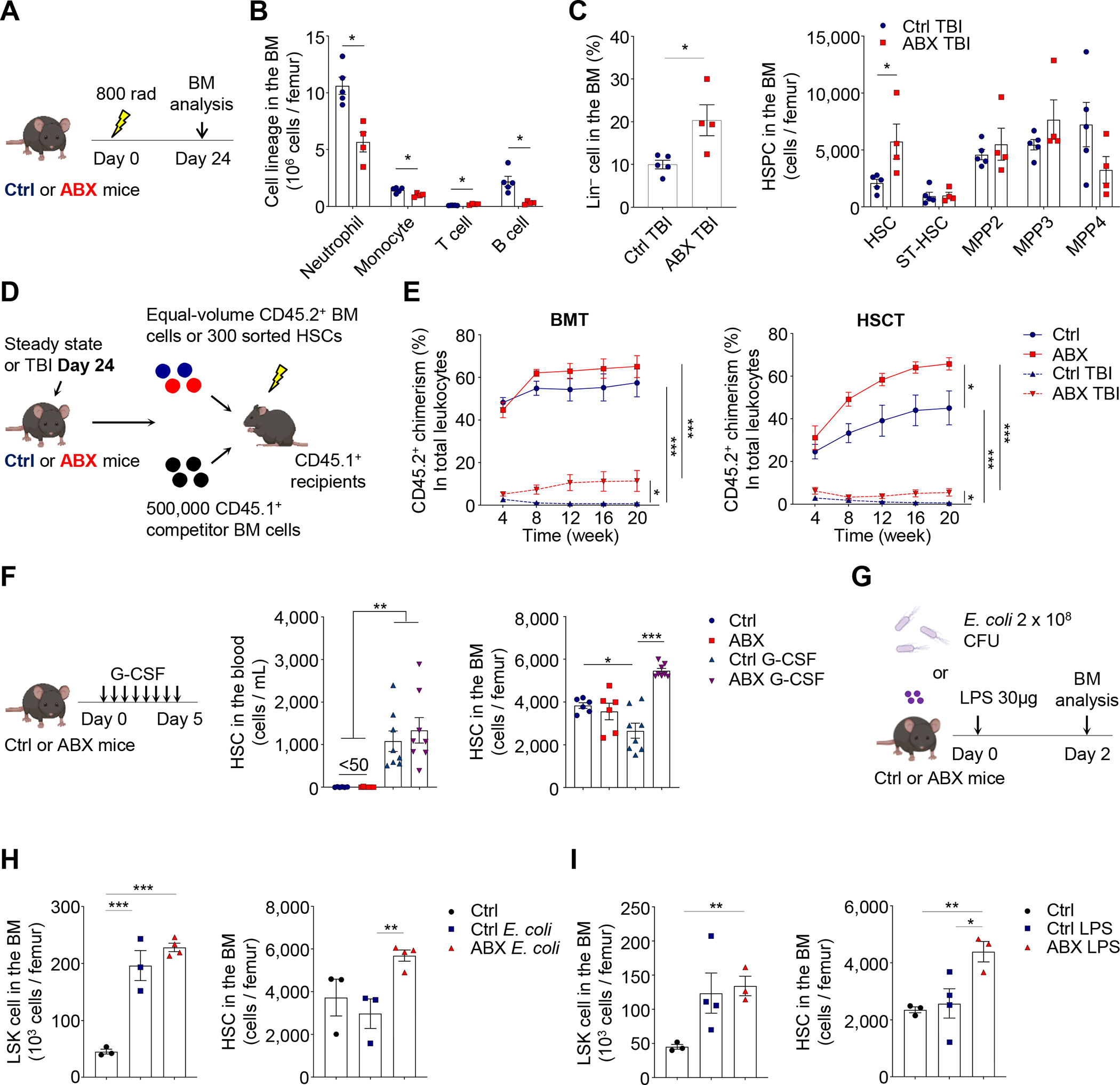
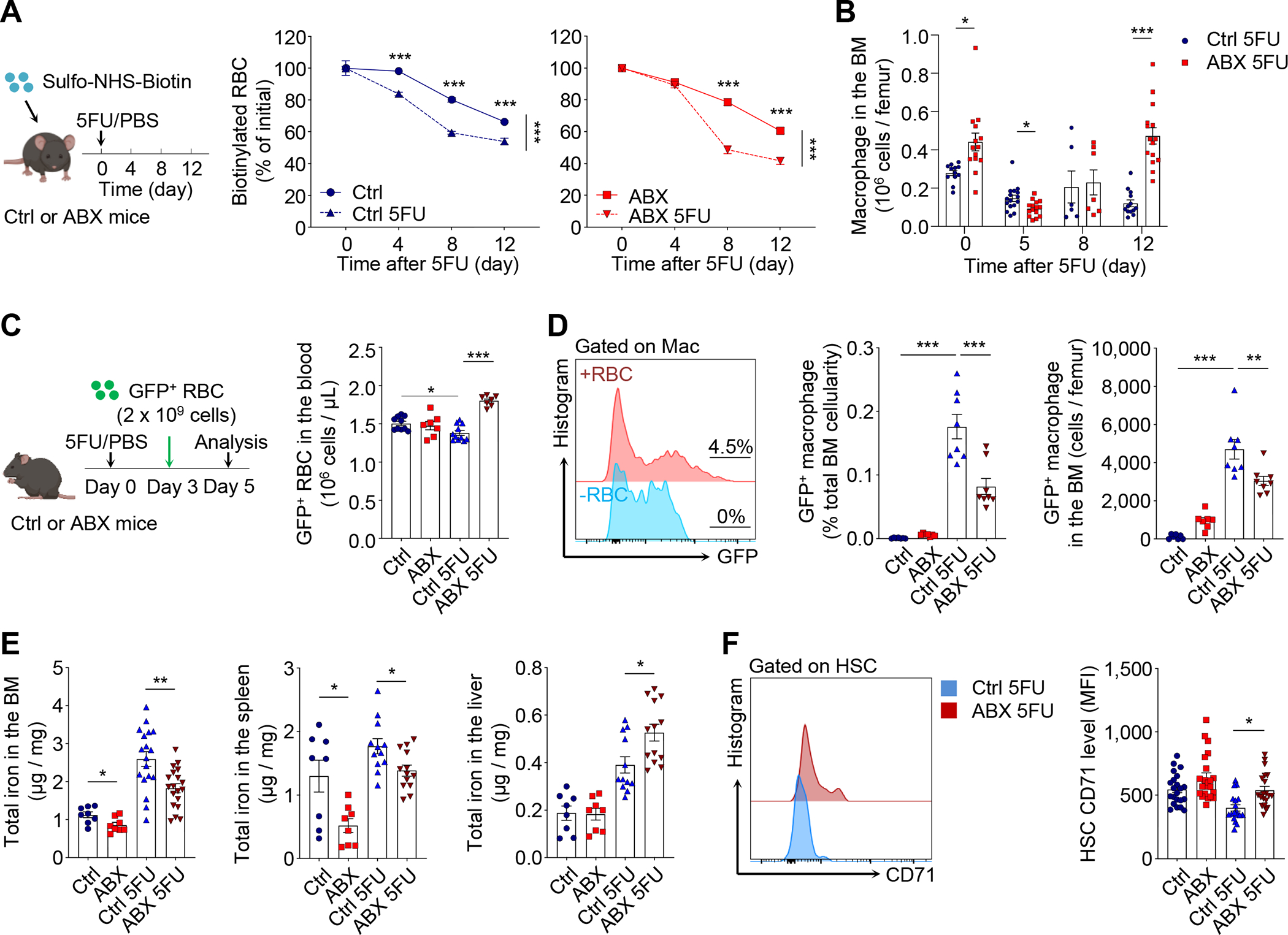
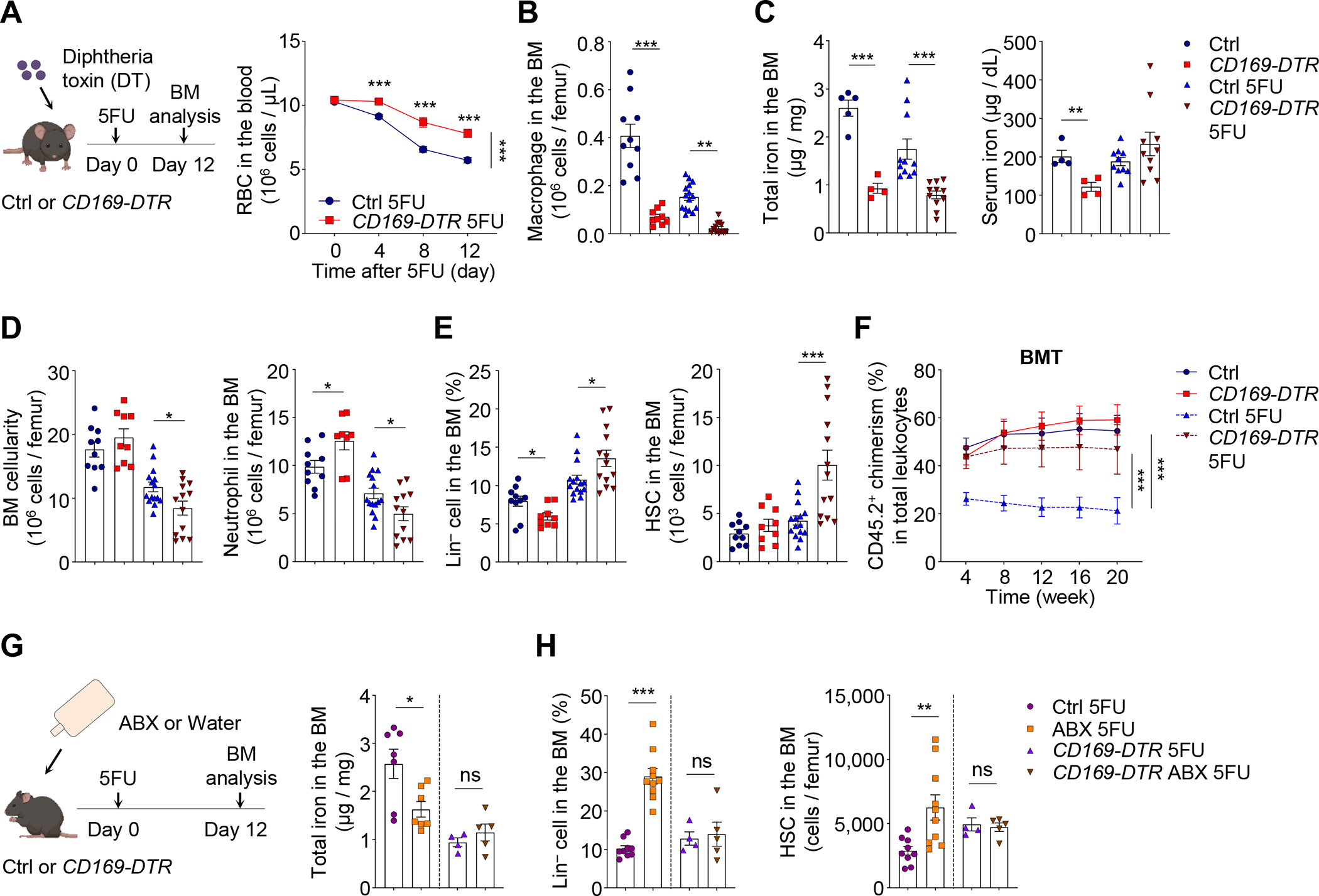
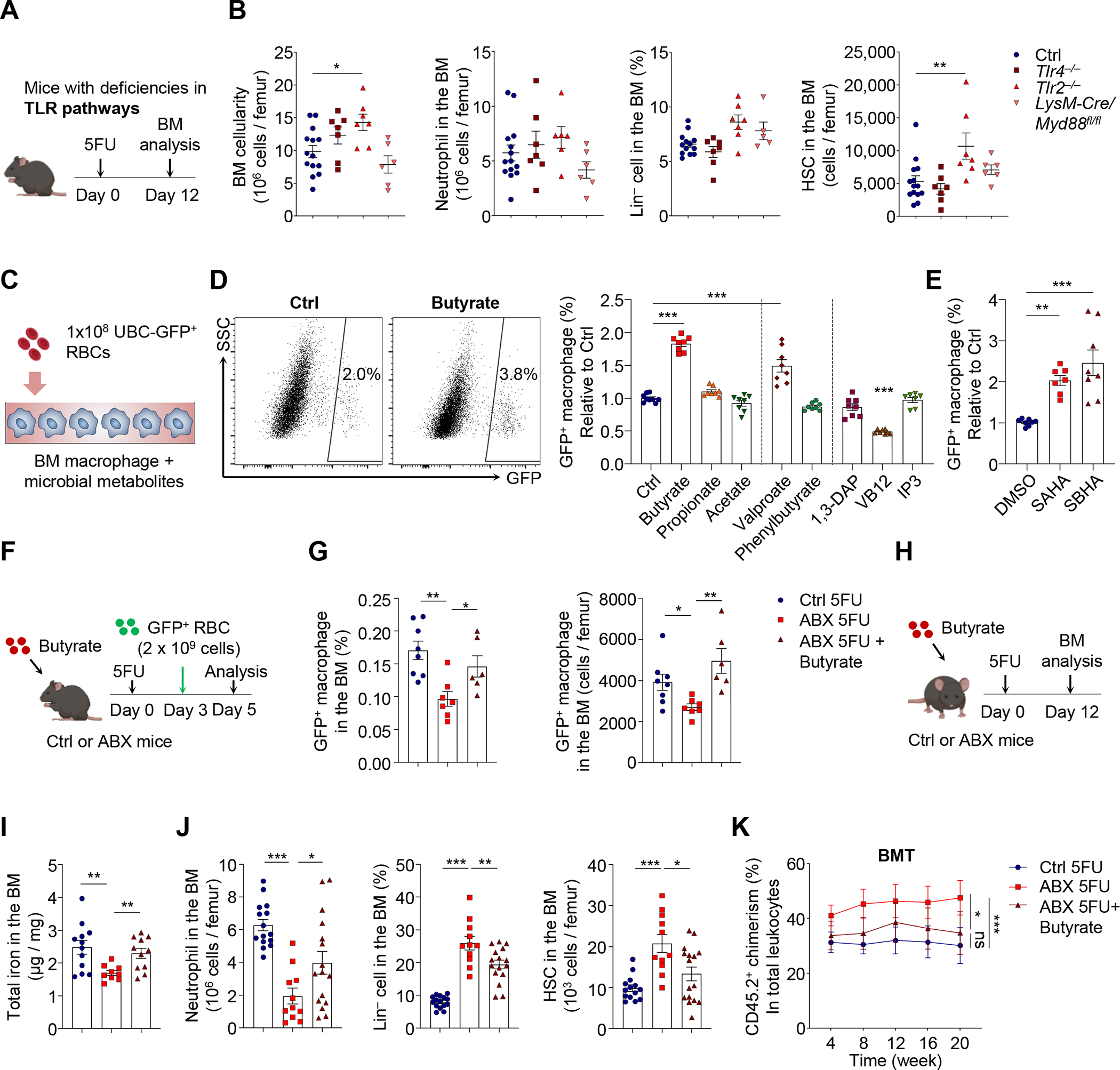
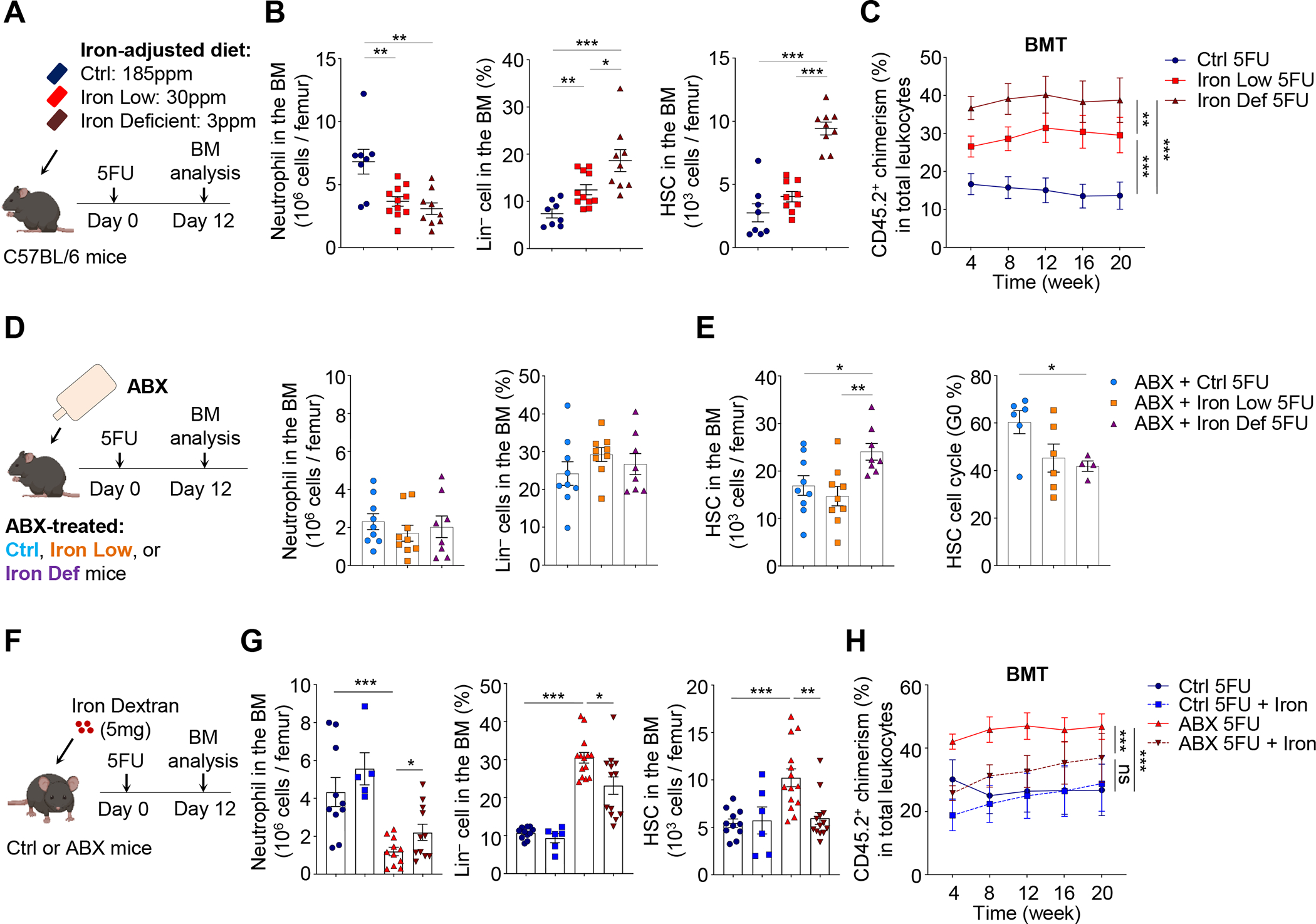
Host microbiota crosstalk is essential for the production and functional modulation of blood-cell lineages. Whether, and if so how, the microbiota influences hematopoietic stem cells (HSCs) is unclear. Here, we show that the microbiota regulates HSC self-renewal and differentiation under stress conditions by modulating local iron availability in the bone marrow (BM). In microbiota-depleted mice, HSC self-renewal was enhanced during regeneration, while the commitment toward differentiation was dramatically compromised. Mechanistically, microbiota depletion selectively impaired the recycling of red blood cells (RBCs) by BM macrophages, resulting in reduced local iron levels without affecting systemic iron homeostasis. Limiting iron availability in food (in vivo) or in culture (ex vivo), or by CD169+ macrophage depletion, enhanced HSC self-renewal and expansion. These results reveal an intricate interplay between the microbiota, macrophages, and iron, and their essential roles in regulating critical HSC fate decisions under stress.
Keywords: erythrophagocytosis; fate decision; hematopoietic regeneration; hematopoietic stem cell; iron; macrophage; microbiota; self-renewal.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

Comment in
-
A microbiome-macrophage-iron axis guides stressed hematopoietic stem cell fate.Cell Stem Cell. 2022 Feb 3;29(2):177-179. doi: 10.1016/j.stem.2022.01.009. Cell Stem Cell. 2022. PMID: 35120614
References
-
- Bacchetti De Gregoris T, Aldred N, Clare AS, and Burgess JG (2011). Improvement of phylum- and class-specific primers for real-time PCR quantification of bacterial taxa. J Microbiol Methods 86, 351–356. - PubMed
-
- Balmer ML, Schurch CM, Saito Y, Geuking MB, Li H, Cuenca M, Kovtonyuk LV, McCoy KD, Hapfelmeier S, Ochsenbein AF, et al. (2014). Microbiota-derived compounds drive steady-state granulopoiesis via MyD88/TICAM signaling. J Immunol 193, 5273–5283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
