

Unique molecular and functional features of extramedullary hematopoietic stem and progenitor cell reservoirs in humans
- PMID: 35073399
- PMCID: PMC7612845
- DOI: 10.1182/blood.2021013450
Unique molecular and functional features of extramedullary hematopoietic stem and progenitor cell reservoirs in humans
Abstract
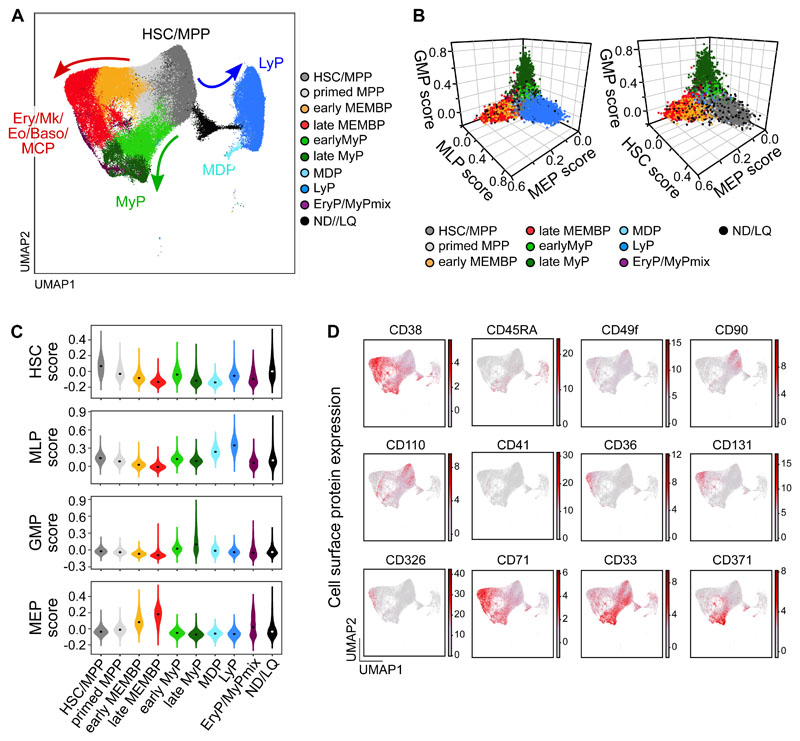
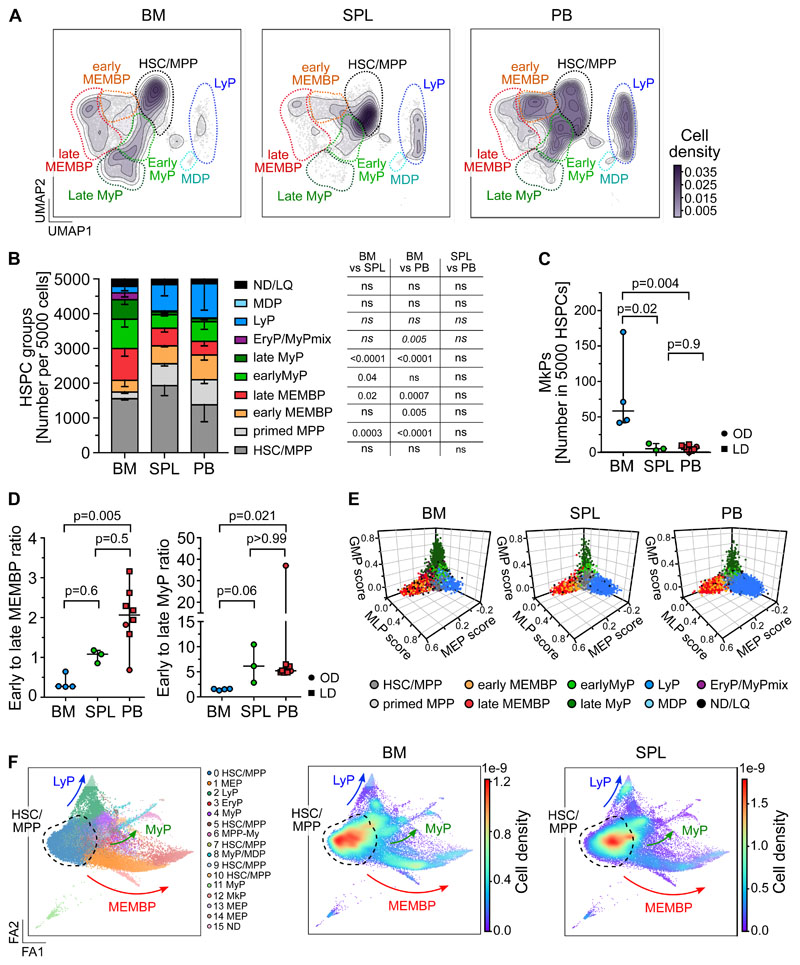
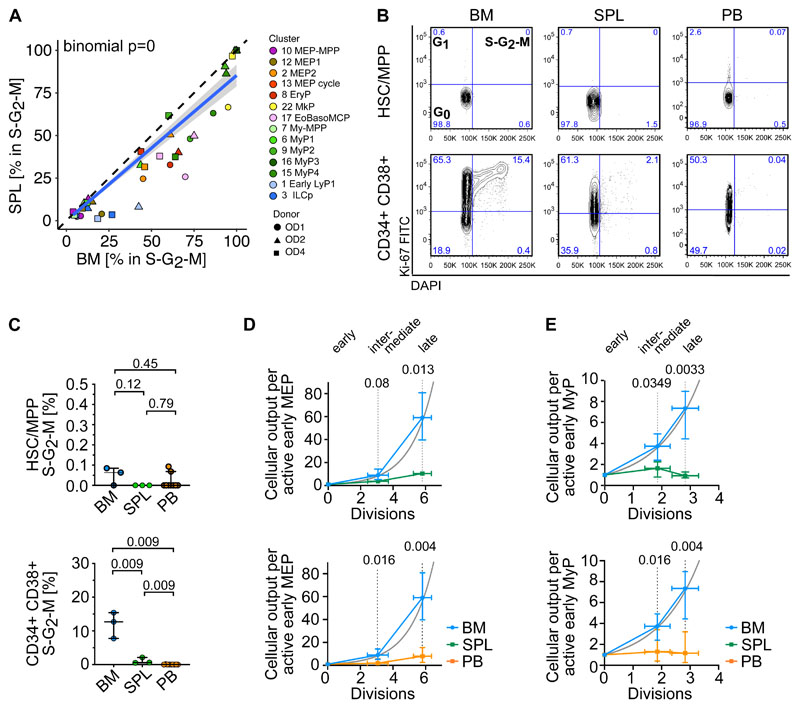
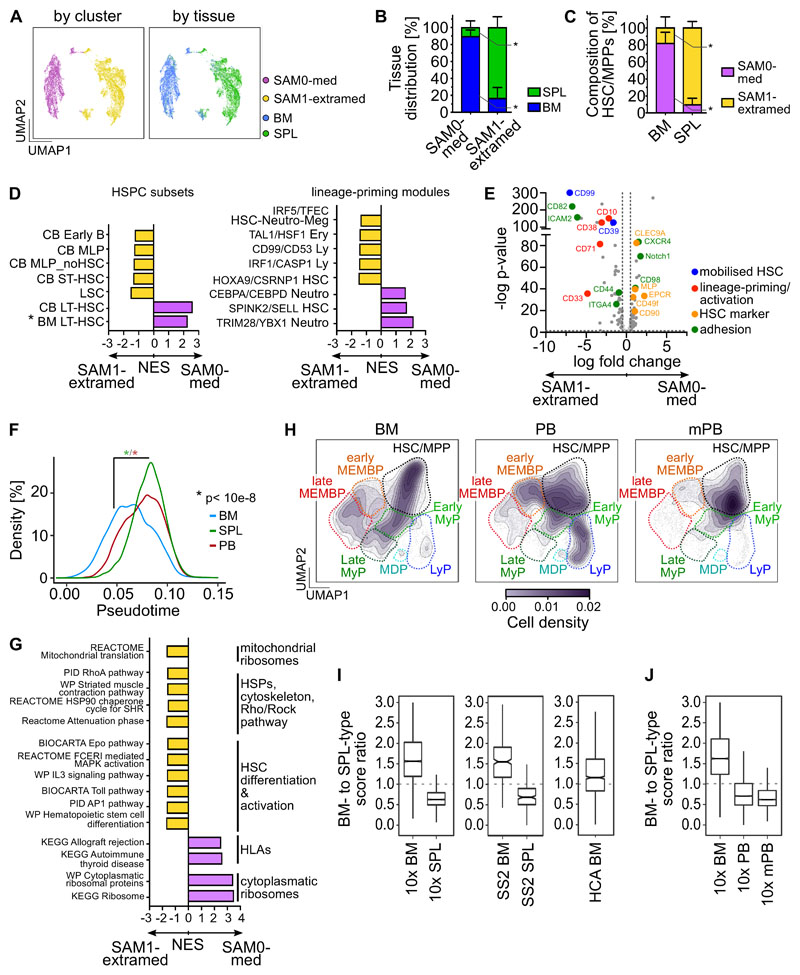
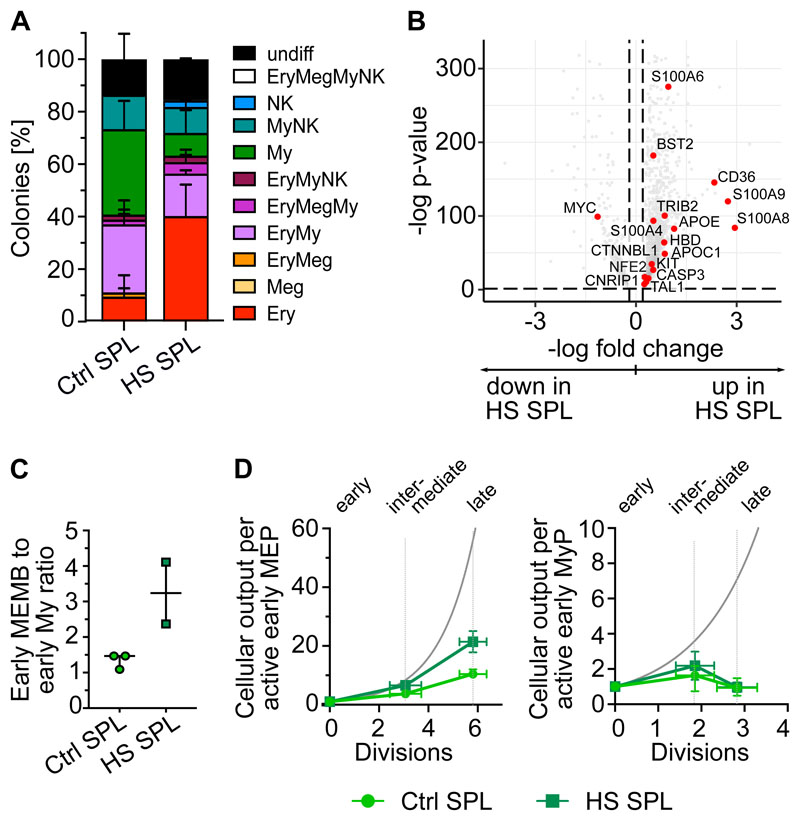
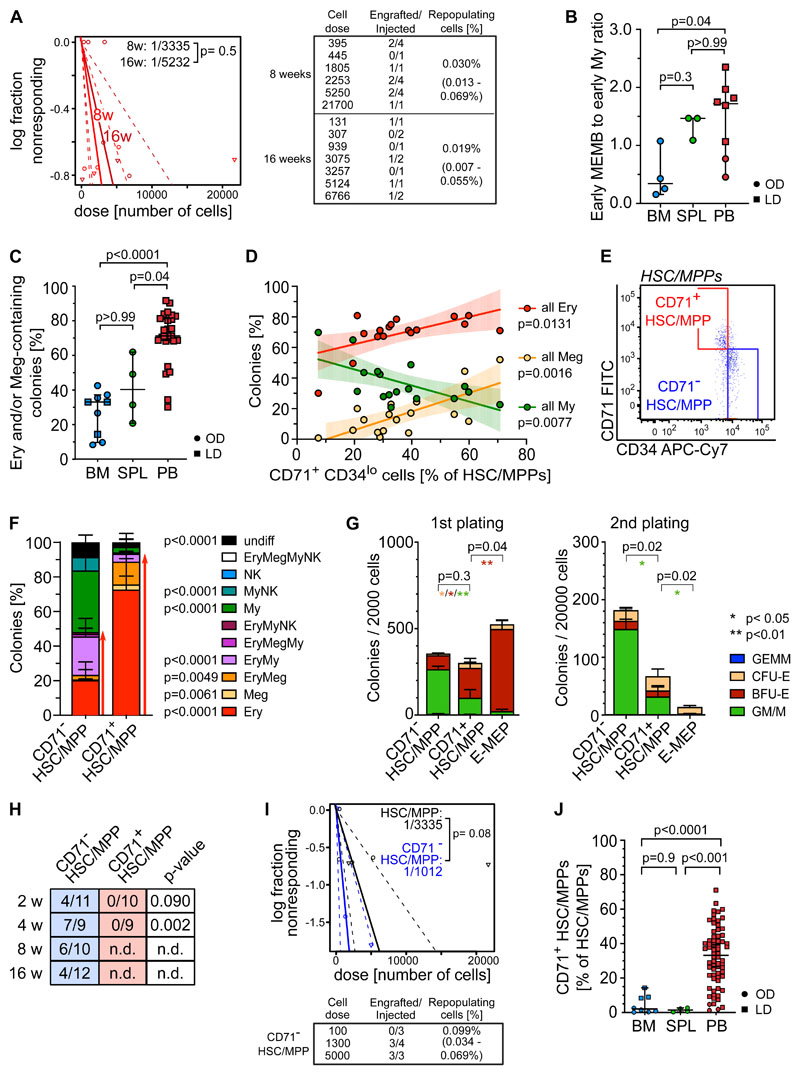
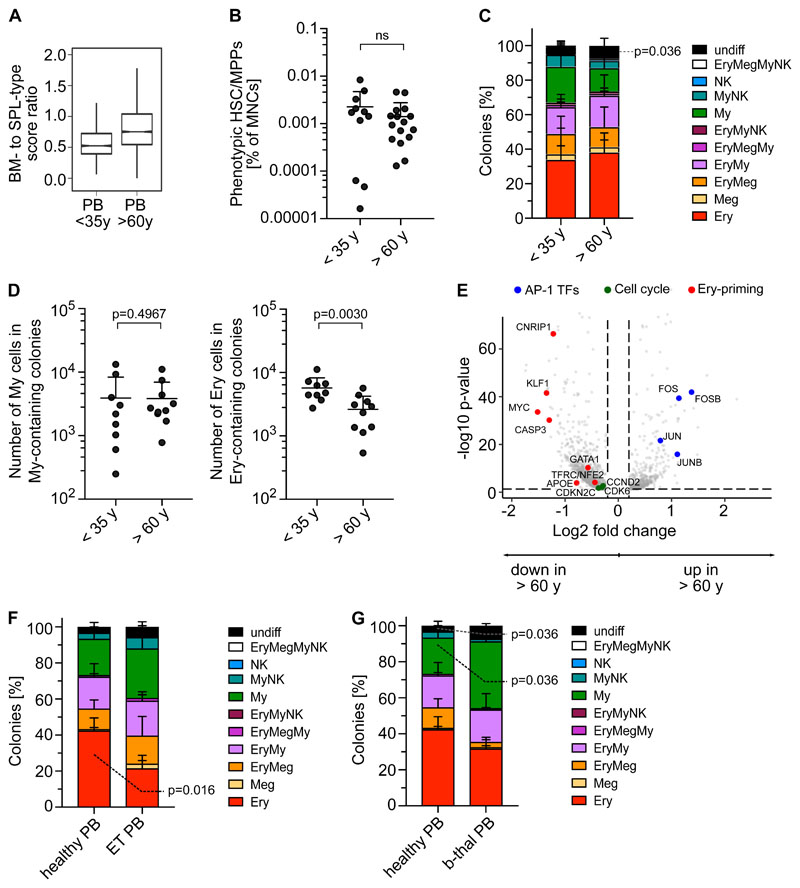
Rare hematopoietic stem and progenitor cell (HSPC) pools outside the bone marrow (BM) contribute to blood production in stress and disease but remain ill-defined. Although nonmobilized peripheral blood (PB) is routinely sampled for clinical management, the diagnosis and monitoring potential of PB HSPCs remain untapped, as no healthy PB HSPC baseline has been reported. Here we comprehensively delineate human extramedullary HSPC compartments comparing spleen, PB, and mobilized PB to BM using single-cell RNA-sequencing and/or functional assays. We uncovered HSPC features shared by extramedullary tissues and others unique to PB. First, in contrast to actively dividing BM HSPCs, we found no evidence of substantial ongoing hematopoiesis in extramedullary tissues at steady state but report increased splenic HSPC proliferative output during stress erythropoiesis. Second, extramedullary hematopoietic stem cells/multipotent progenitors (HSCs/MPPs) from spleen, PB, and mobilized PB share a common transcriptional signature and increased abundance of lineage-primed subsets compared with BM. Third, healthy PB HSPCs display a unique bias toward erythroid-megakaryocytic differentiation. At the HSC/MPP level, this is functionally imparted by a subset of phenotypic CD71+ HSCs/MPPs, exclusively producing erythrocytes and megakaryocytes, highly abundant in PB but rare in other adult tissues. Finally, the unique erythroid-megakaryocytic-skewing of PB is perturbed with age in essential thrombocythemia and β-thalassemia. Collectively, we identify extramedullary lineage-primed HSPC reservoirs that are nonproliferative in situ and report involvement of splenic HSPCs during demand-adapted hematopoiesis. Our data also establish aberrant composition and function of circulating HSPCs as potential clinical indicators of BM dysfunction.
© 2022 by The American Society of Hematology.
Conflict of interest statement
No conflicts of interest to declare
Figures
Comment in
-
Extramedullary hematopoietic stem cells.Blood. 2022 Jun 9;139(23):3353-3354. doi: 10.1182/blood.2022015879. Blood. 2022. PMID: 35679074 No abstract available.
References
-
- Soderdahl G, Tammik C, Remberger M, Ringden O. Cadaveric bone marrow and spleen cells for transplantation. Bone Marrow Transplantation. 1998;21:79–84. - PubMed
-
- Wright DE, Wagers AJ, Gulati AP, Johnson FL, Weissman IL. Physiological Migration of Hematopoietic Stem and Progenitor Cells. Science. 2001;294(5548):1933–1936. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
