

IQGAP1 scaffolding links phosphoinositide kinases to cytoskeletal reorganization
- PMID: 35077666
- PMCID: PMC8943696
- DOI: 10.1016/j.bpj.2022.01.018
IQGAP1 scaffolding links phosphoinositide kinases to cytoskeletal reorganization
Abstract
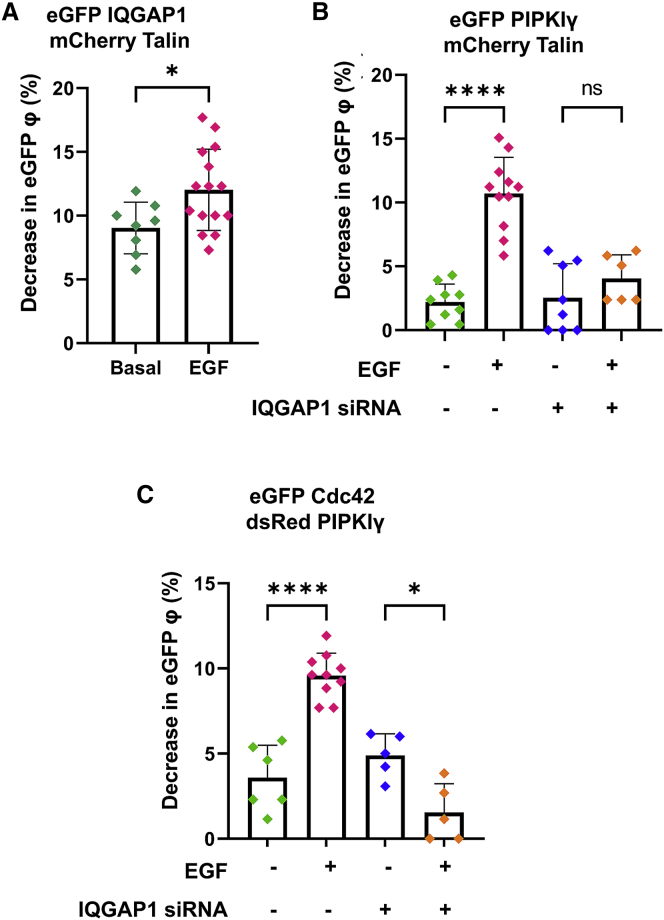
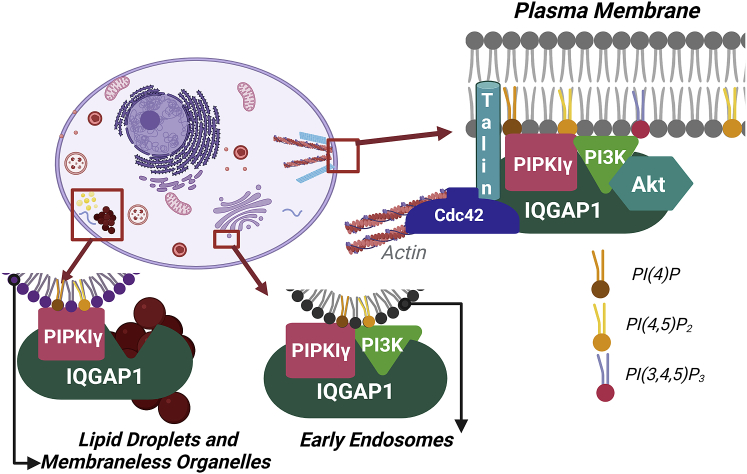
IQGAP1 is a multidomain scaffold protein that coordinates the direction and impact of multiple signaling pathways by scaffolding its various binding partners. However, the spatial and temporal resolution of IQGAP1 scaffolding remains unclear. Here, we use fluorescence imaging and correlation methods that allow for real-time live-cell changes in IQGAP1 localization and complex formation during signaling. We find that IQGAP1 and PIPKIγ interact on both the plasma membrane and in cytosol. Epidermal growth factor (EGF) stimulation, which can initiate cytoskeletal changes, drives the movement of the cytosolic pool toward the plasma membrane to promote cytoskeletal changes. We also observe that a significant population of cytosolic IQGAP1-PIPKIγ complexes localize to early endosomes, and in some instances form aggregated clusters which become highly mobile upon EGF stimulation. Our imaging studies show that PIPKIγ and PI3K bind simultaneously to IQGAP1, which may accelerate conversion of PI4P to PI(3,4,5)P3 that is required for cytoskeletal changes. Additionally, we find that IQGAP1 is responsible for PIPKIγ association with two proteins associated with cytoskeletal changes, talin and Cdc42, during EGF stimulation. These results directly show that IQGAP1 provides a physical link between phosphoinositides (through PIPKIγ), focal adhesion formation (through talin), and cytoskeletal reorganization (through Cdc42) upon EGF stimulation. Taken together, our results support the importance of IQGAP1 in regulating cell migration by linking phosphoinositide lipid signaling with cytoskeletal reorganization.
Copyright © 2022 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
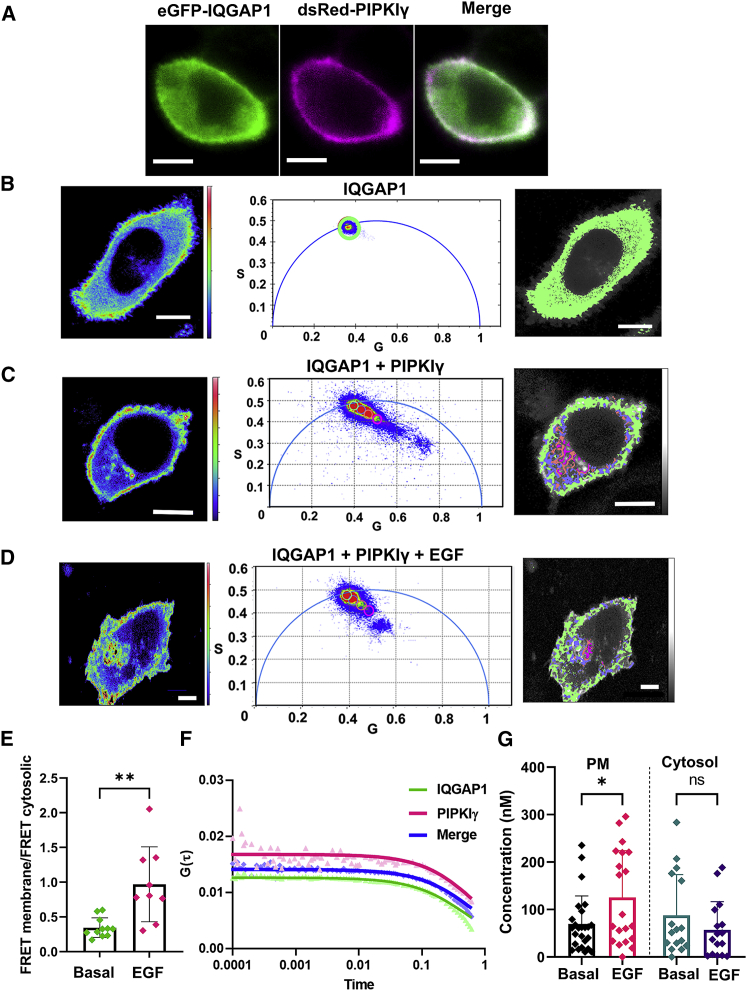
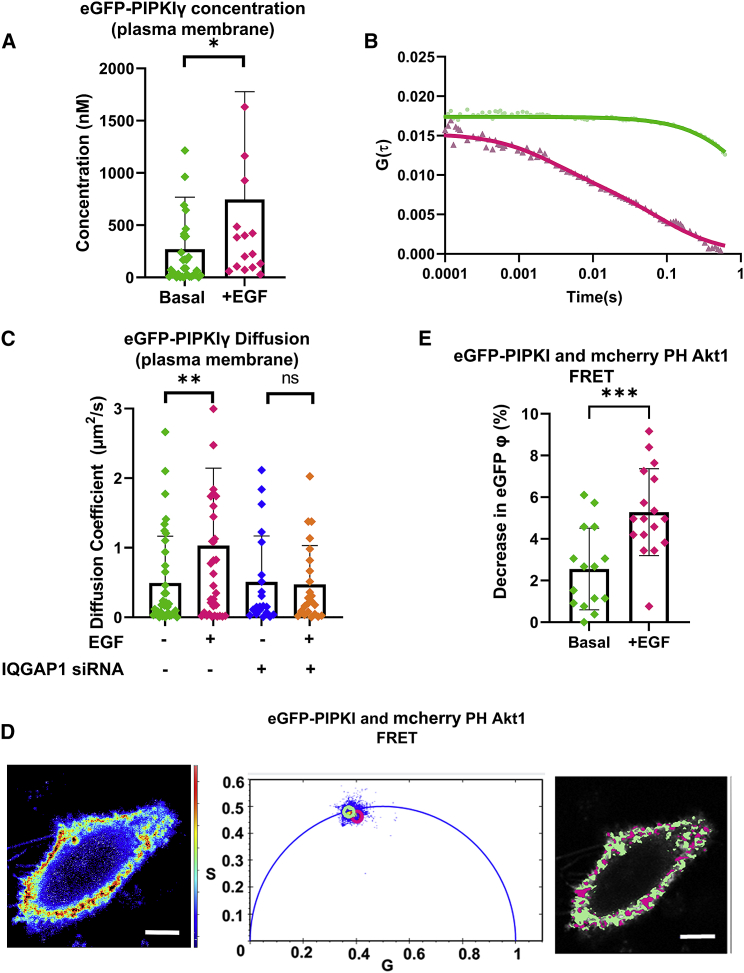
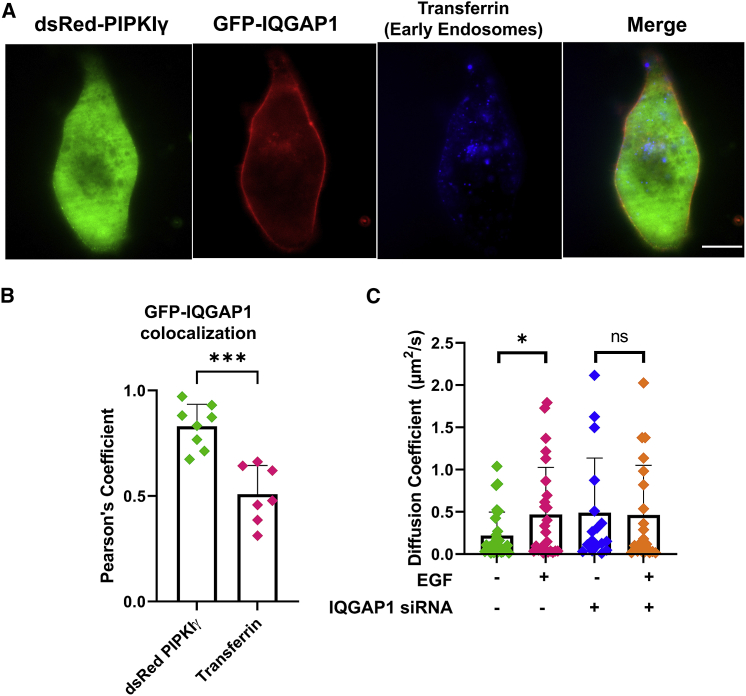
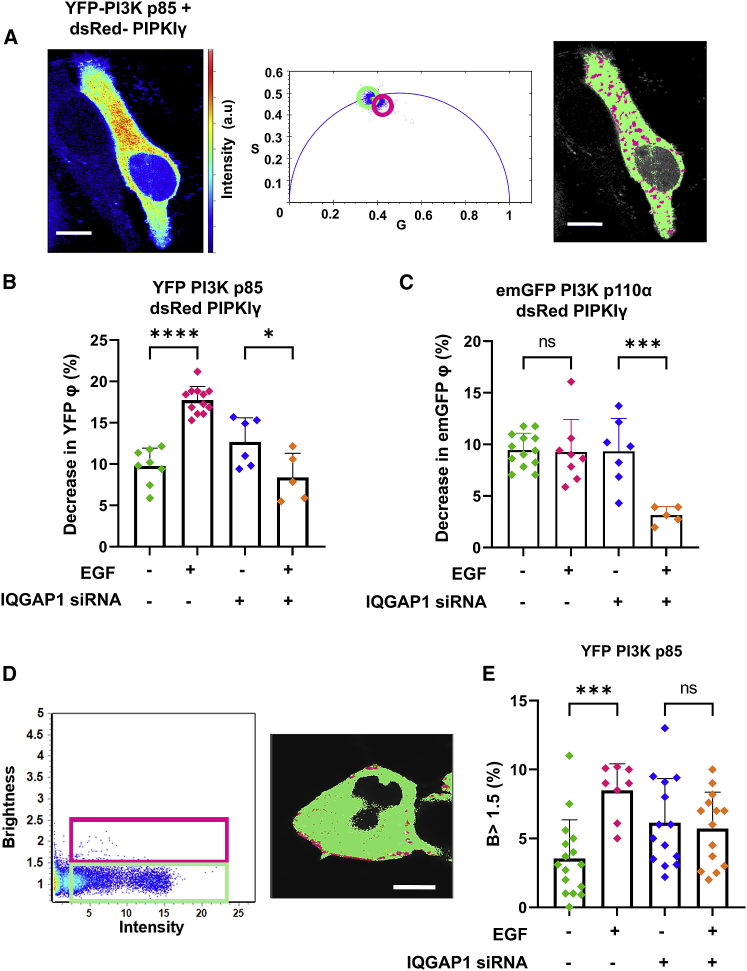
References
-
- Berridge M.J., Irvine R.F. Inositol trisphosphate, a novel second messenger in cellular signal transduction. Nature. 1984;312:315–321. - PubMed
-
- Prestwich G.D. Phosphoinositide signaling; from affinity probes to pharmaceutical targets. Chem. Biol. 2004;11:619–637. - PubMed
-
- Bunney T.D., Katan M. Phosphoinositide signalling in cancer: beyond PI3K and PTEN. Nat. Rev. Cancer. 2010;10:342–352. - PubMed
-
- Park S.J., Itoh T., Takenawa T. Phosphatidylinositol 4-phosphate 5-kinase type I is regulated through phosphorylation response by extracellular stimuli. J. Biol. Chem. 2001;276:4781–4787. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
