

The (Lack of) DNA Double-Strand Break Repair Pathway Choice During V(D)J Recombination
- PMID: 35082840
- PMCID: PMC8785701
- DOI: 10.3389/fgene.2021.823943
The (Lack of) DNA Double-Strand Break Repair Pathway Choice During V(D)J Recombination
Abstract
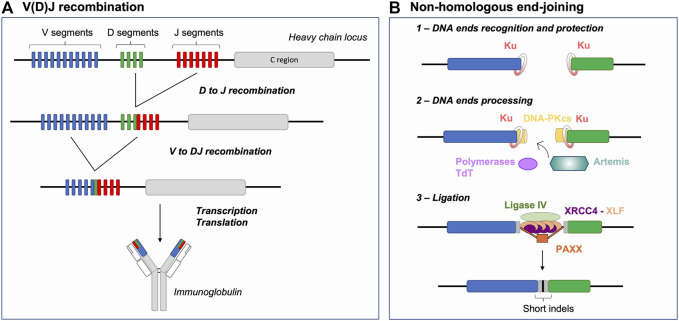
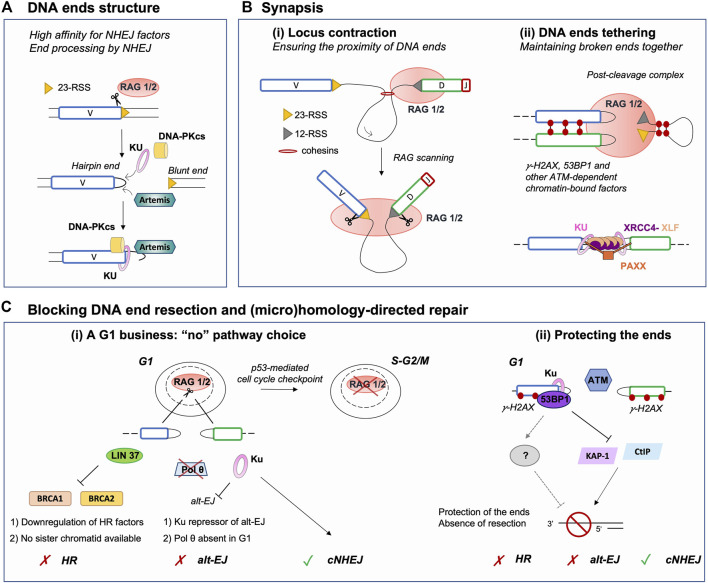
DNA double-strand breaks (DSBs) are highly toxic lesions that can be mended via several DNA repair pathways. Multiple factors can influence the choice and the restrictiveness of repair towards a given pathway in order to warrant the maintenance of genome integrity. During V(D)J recombination, RAG-induced DSBs are (almost) exclusively repaired by the non-homologous end-joining (NHEJ) pathway for the benefit of antigen receptor gene diversity. Here, we review the various parameters that constrain repair of RAG-generated DSBs to NHEJ, including the peculiarity of DNA DSB ends generated by the RAG nuclease, the establishment and maintenance of a post-cleavage synaptic complex, and the protection of DNA ends against resection and (micro)homology-directed repair. In this physiological context, we highlight that certain DSBs have limited DNA repair pathway choice options.
Keywords: DNA double-strand break; DNA double-strand break repair pathway choice; DNA end resection; V(D)J recombination; homology-directed repair; non-homologous end-joining.
Copyright © 2022 Libri, Marton and Deriano.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources