

Amplification of human interneuron progenitors promotes brain tumors and neurological defects
- PMID: 35084981
- PMCID: PMC7613689
- DOI: 10.1126/science.abf5546
Amplification of human interneuron progenitors promotes brain tumors and neurological defects
Abstract
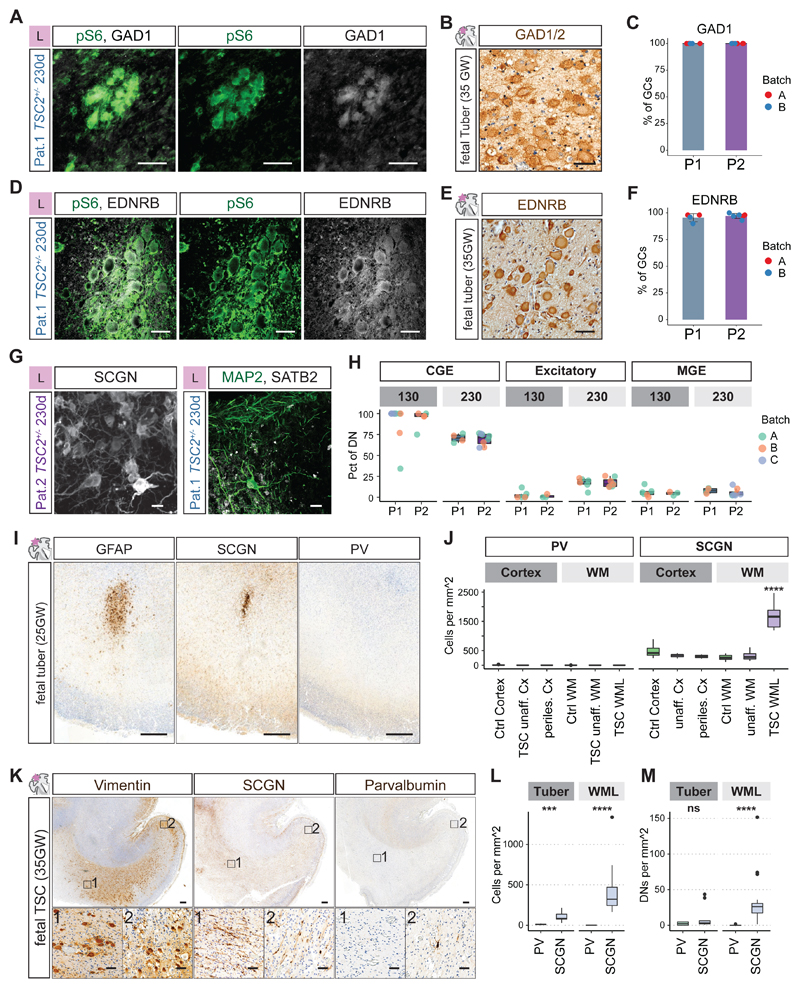
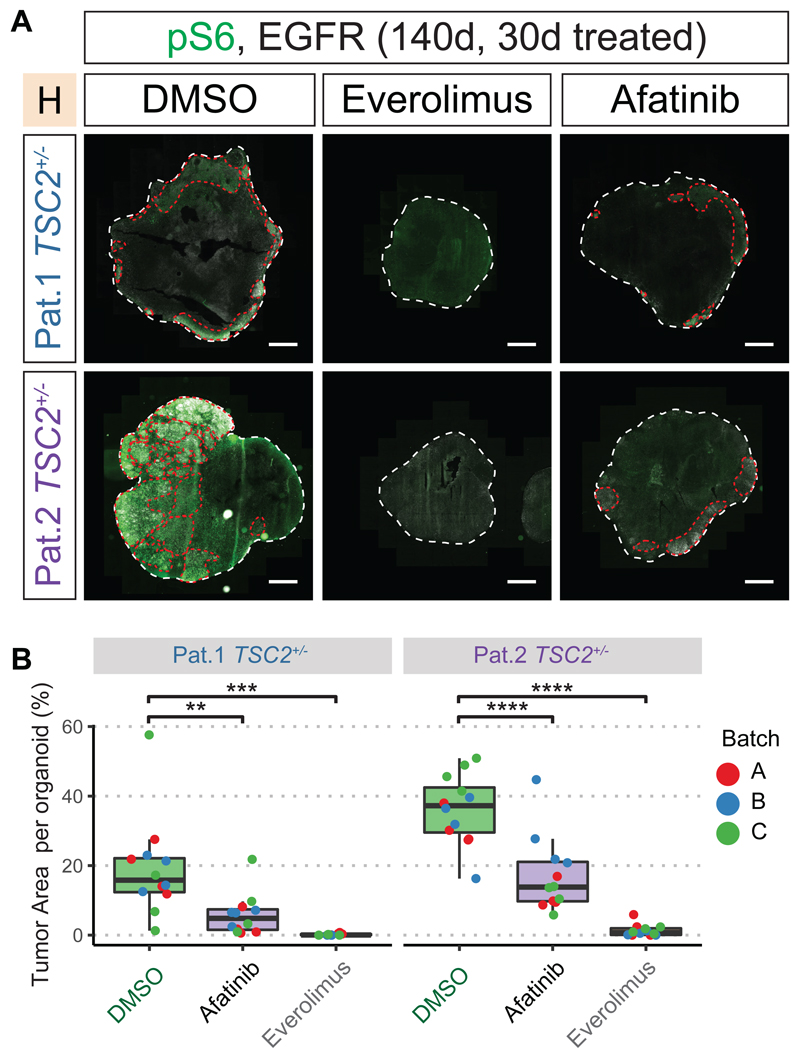
Evolutionary development of the human brain is characterized by the expansion of various brain regions. Here, we show that developmental processes specific to humans are responsible for malformations of cortical development (MCDs), which result in developmental delay and epilepsy in children. We generated a human cerebral organoid model for tuberous sclerosis complex (TSC) and identified a specific neural stem cell type, caudal late interneuron progenitor (CLIP) cells. In TSC, CLIP cells over-proliferate, generating excessive interneurons, brain tumors, and cortical malformations. Epidermal growth factor receptor inhibition reduces tumor burden, identifying potential treatment options for TSC and related disorders. The identification of CLIP cells reveals the extended interneuron generation in the human brain as a vulnerability for disease. In addition, this work demonstrates that analyzing MCDs can reveal fundamental insights into human-specific aspects of brain development.
Conflict of interest statement
Figures
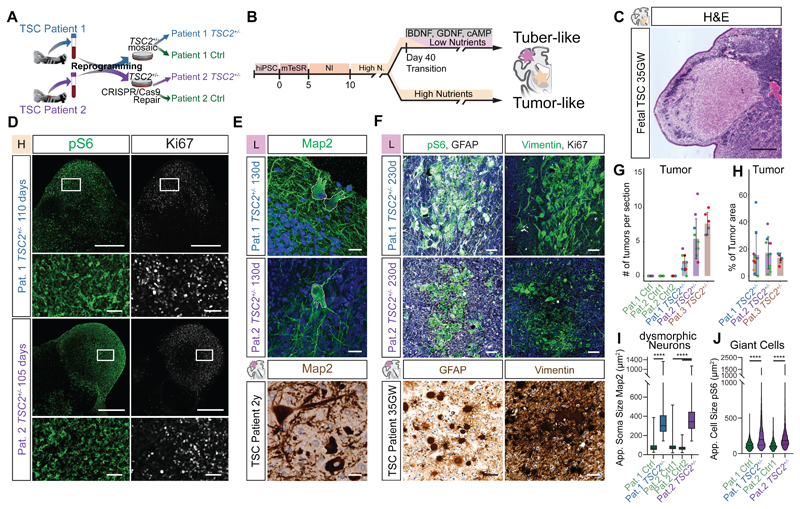
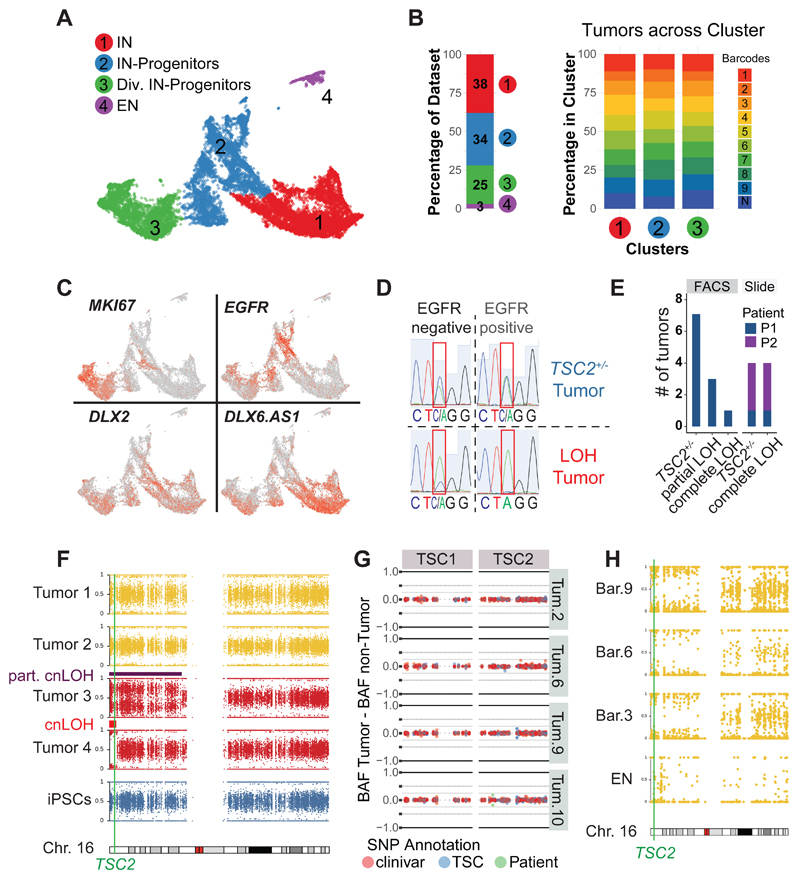
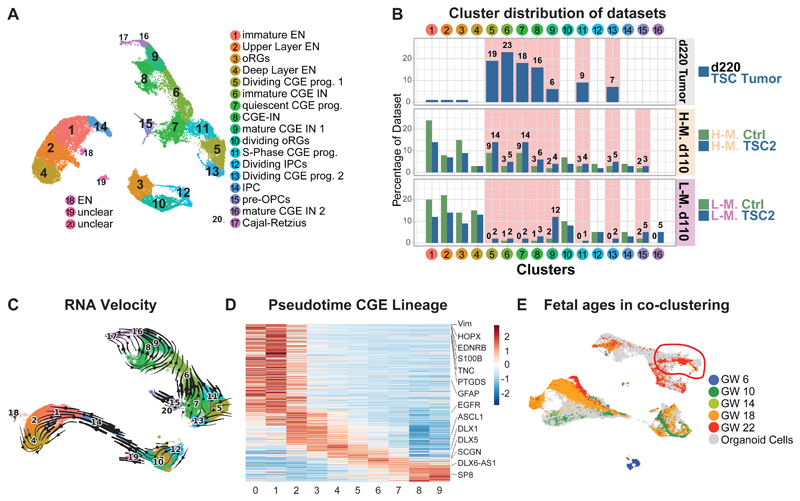
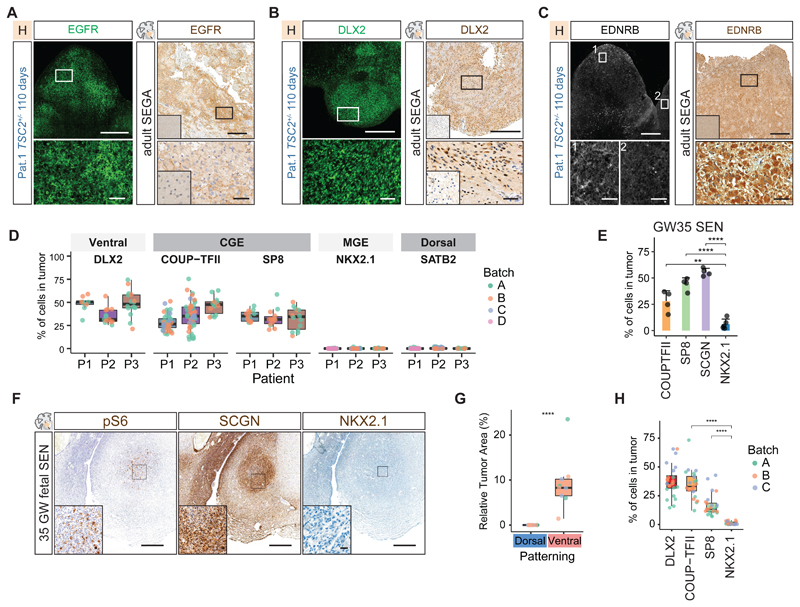
Comment in
-
Modeling tuberous sclerosis with organoids.Science. 2022 Jan 28;375(6579):382-383. doi: 10.1126/science.abn6158. Epub 2022 Jan 27. Science. 2022. PMID: 35084978
-
Tubers and Tumors Are CLIPped Together in Tuberous Sclerosis Complex.Epilepsy Curr. 2022 May 10;22(4):255-257. doi: 10.1177/15357597221101278. eCollection 2022 Jul-Aug. Epilepsy Curr. 2022. PMID: 36187152 Free PMC article. No abstract available.
References
-
- Kuzniecky RI. MRI in cerebral developmental malformations and epilepsy. Magnetic Resonance Imaging. 1995;13:1137–1145. - PubMed
-
- Thiele EA. Managing and understanding epilepsy in tuberous sclerosis complex. Epilepsia. 2010;51(Suppl 1):90–91. - PubMed
-
- Ruppe V, et al. Developmental brain abnormalities in tuberous sclerosis complex: a comparative tissue analysis of cortical tubers and perituberal cortex. Epilepsia. 2014;55:539–550. - PubMed
-
- Henske EP, Jozwiak S, Kingswood JC, Sampson JR, Thiele EA. Tuberous sclerosis complex. Nat Rev Dis Primers. 2016;2:16035. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
