

Parallel evolution of a splicing program controlling neuronal excitability in flies and mammals
- PMID: 35089784
- PMCID: PMC8797185
- DOI: 10.1126/sciadv.abk0445
Parallel evolution of a splicing program controlling neuronal excitability in flies and mammals
Abstract
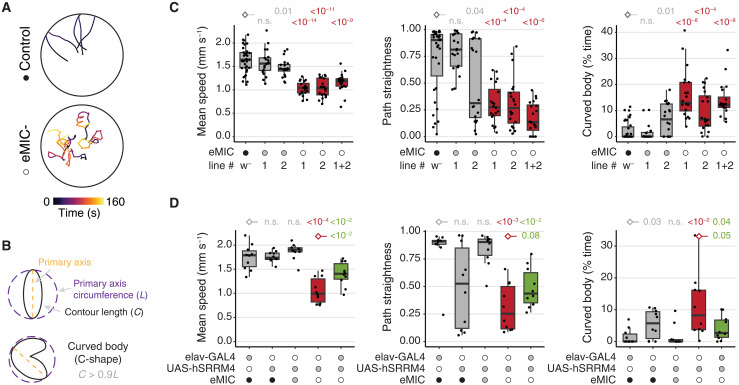
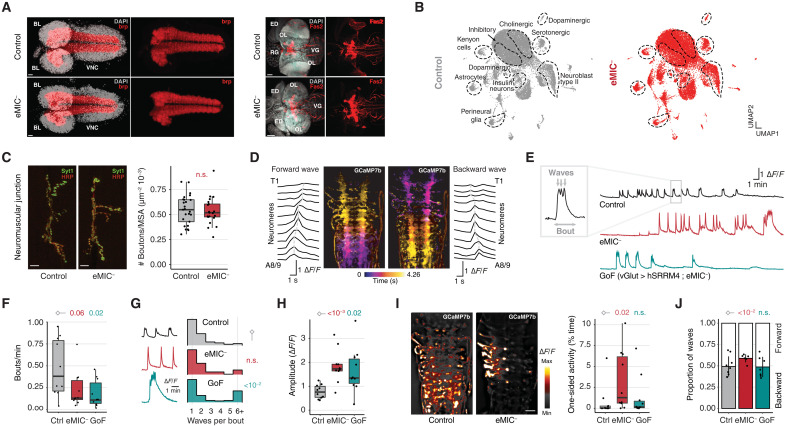
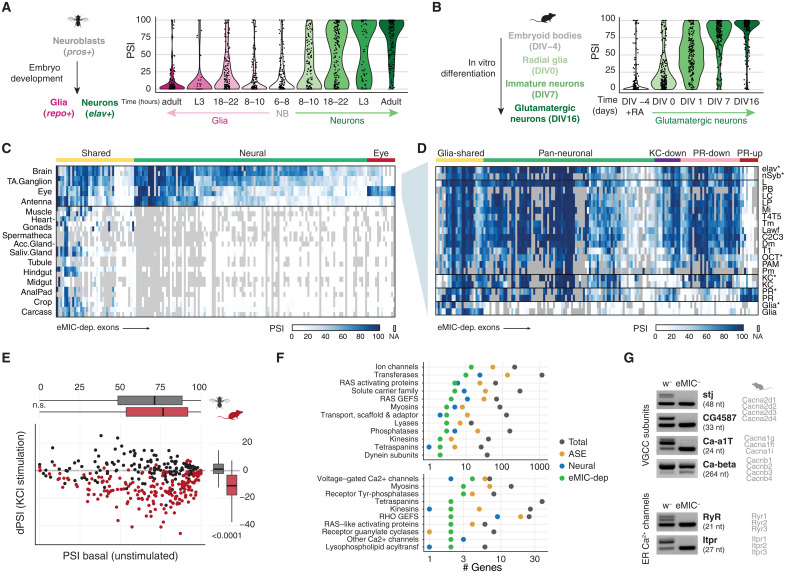
Alternative splicing increases neuronal transcriptomic complexity throughout animal phylogeny. To delve into the mechanisms controlling the assembly and evolution of this regulatory layer, we characterized the neuronal microexon program in Drosophila and compared it with that of mammals. In nonvertebrate bilaterians, this splicing program is restricted to neurons by the posttranscriptional processing of the enhancer of microexons (eMIC) domain in Srrm234. In Drosophila, this processing is dependent on regulation by Elav/Fne. eMIC deficiency or misexpression leads to widespread neurological alterations largely emerging from impaired neuronal activity, as revealed by a combination of neuronal imaging experiments and cell type-specific rescues. These defects are associated with the genome-wide skipping of short neural exons, which are strongly enriched in ion channels. We found no overlap of eMIC-regulated exons between flies and mice, illustrating how ancient posttranscriptional programs can evolve independently in different phyla to affect distinct cellular modules while maintaining cell-type specificity.
Figures






References
-
- Schmucker D., Clemens J. C., Shu H., Worby C. A., Xiao J., Muda M., Dixon J. E., Zipursky S. L., Drosophila Dscam is an axon guidance receptor exhibiting extraordinary molecular diversity. Cell 101, 671–684 (2000). - PubMed
-
- Gilbert W., Why genes in pieces? Nature 271, 501–501 (1978). - PubMed
-
- Barbosa-Morais N. L., Irimia M., Pan Q., Xiong H. Y., Gueroussov S., Lee L. J., Slobodeniuc V., Kutter C., Watt S., Çolak R., Kim T. H., Misquitta-Ali C. M., Wilson M. D., Kim P. M., Odom D. T., Frey B. J., Blencowe B. J., The evolutionary landscape of alternative splicing in vertebrate species. Science 338, 1587–1593 (2012). - PubMed
-
- Brown J. B., Boley N., Eisman R., May G. E., Stoiber M. H., Duff M. O., Booth B. W., Wen J., Park S., Suzuki A. M., Wan K. H., Yu C., Zhang D., Carlson J. W., Cherbas L., Eads B. D., Miller D., Mockaitis K., Roberts J., Davis C. A., Frise E., Hammonds A. S., Olson S., Shenker S., Sturgill D., Samsonova A. A., Weiszmann R., Robinson G., Hernandez J., Andrews J., Bickel P. J., Carninci P., Cherbas P., Gingeras T. R., Hoskins R. A., Kaufman T. C., Lai E. C., Oliver B., Perrimon N., Graveley B. R., Celniker S. E., Diversity and dynamics of the Drosophila transcriptome. Nature 512, 393–399 (2014). - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
