

Development of Tbet- and CD11c-expressing B cells in a viral infection requires T follicular helper cells outside of germinal centers
- PMID: 35090581
- PMCID: PMC8965751
- DOI: 10.1016/j.immuni.2022.01.002
Development of Tbet- and CD11c-expressing B cells in a viral infection requires T follicular helper cells outside of germinal centers
Abstract
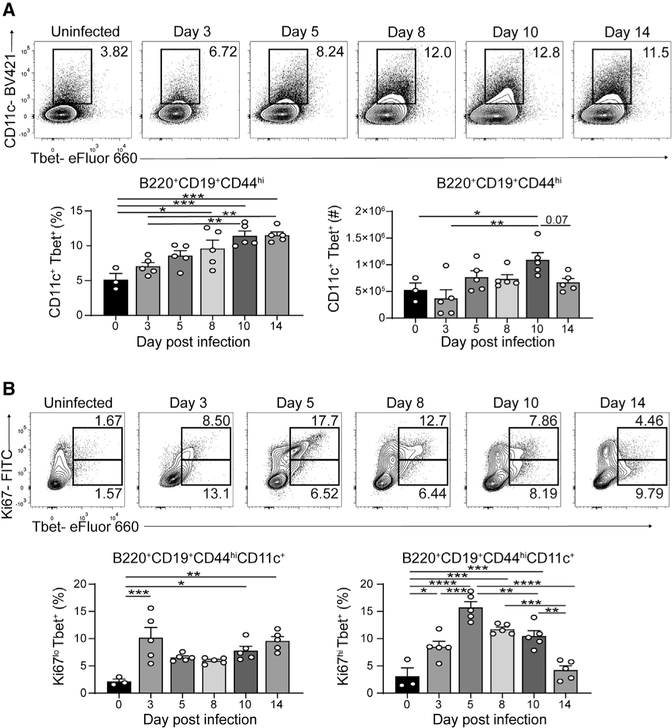
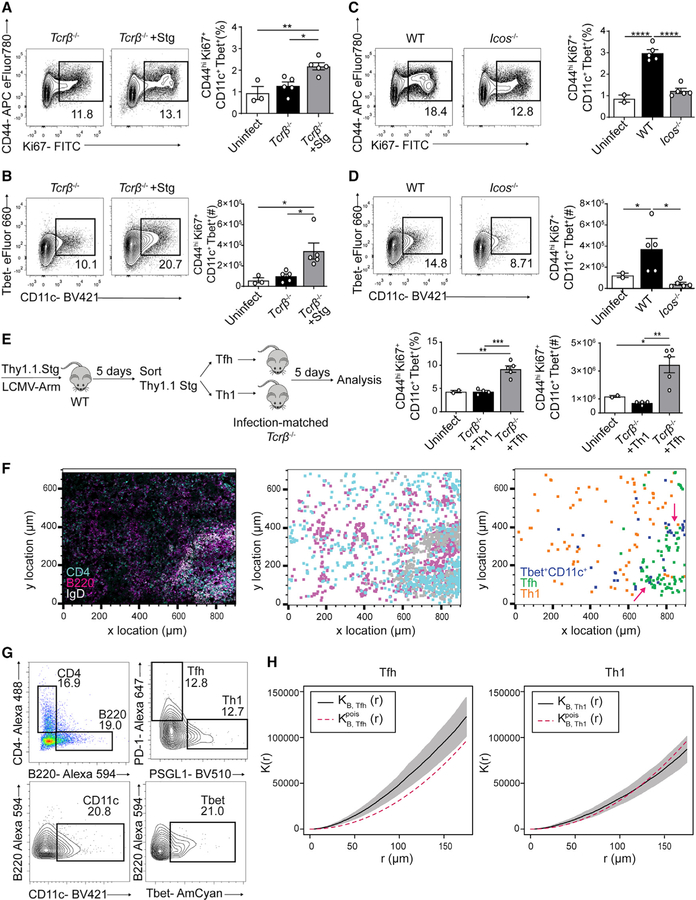
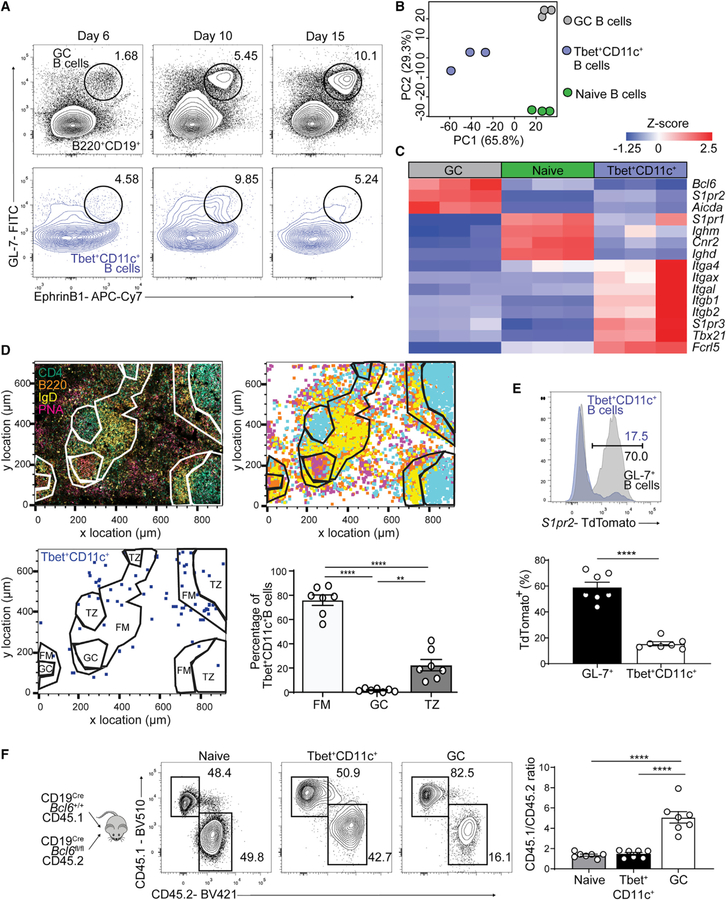
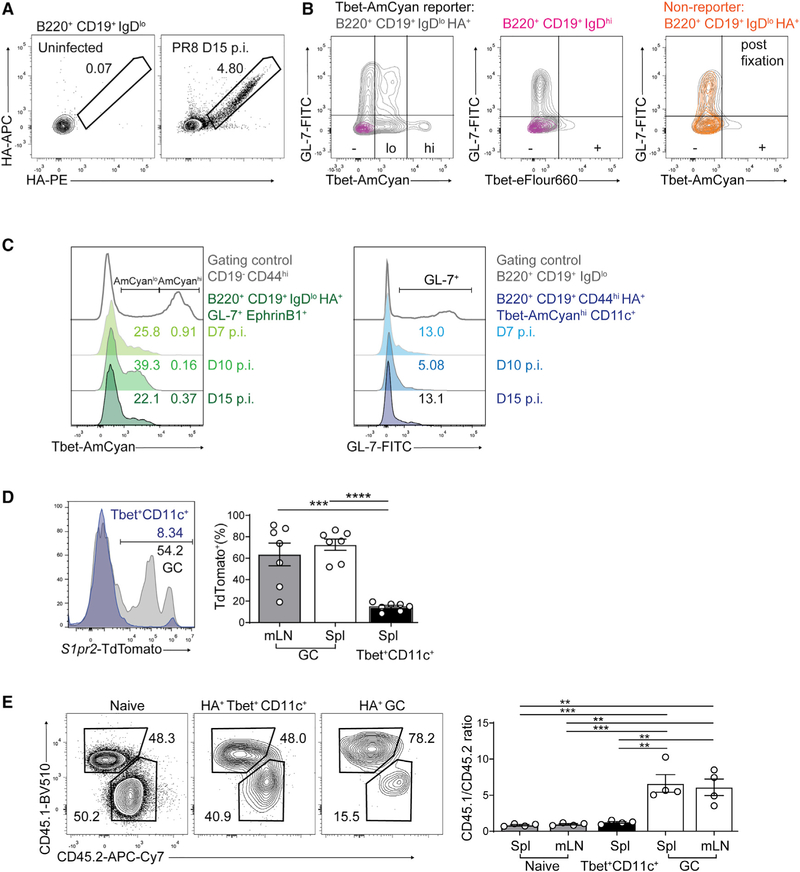
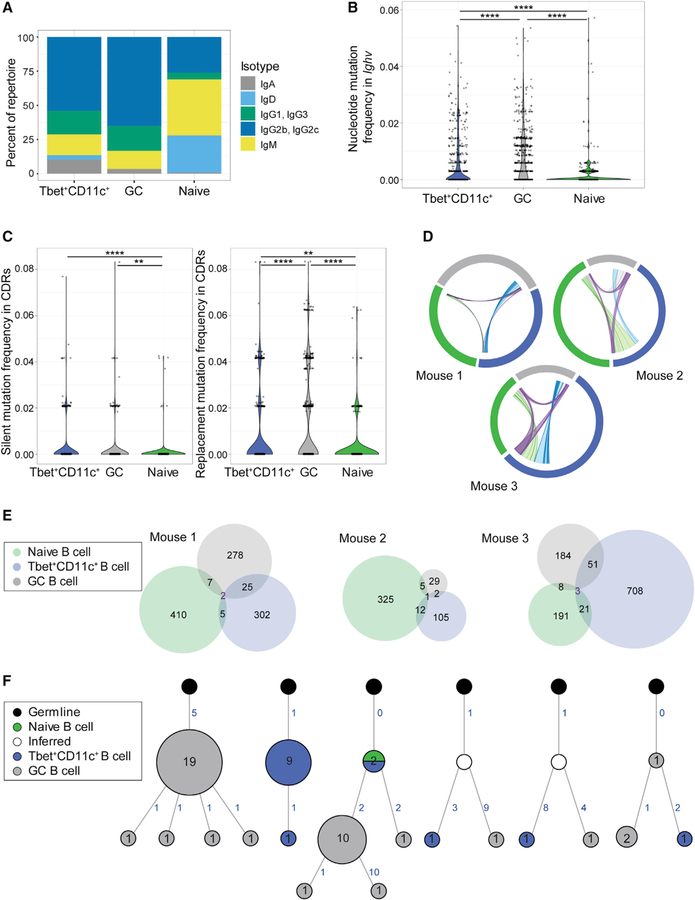
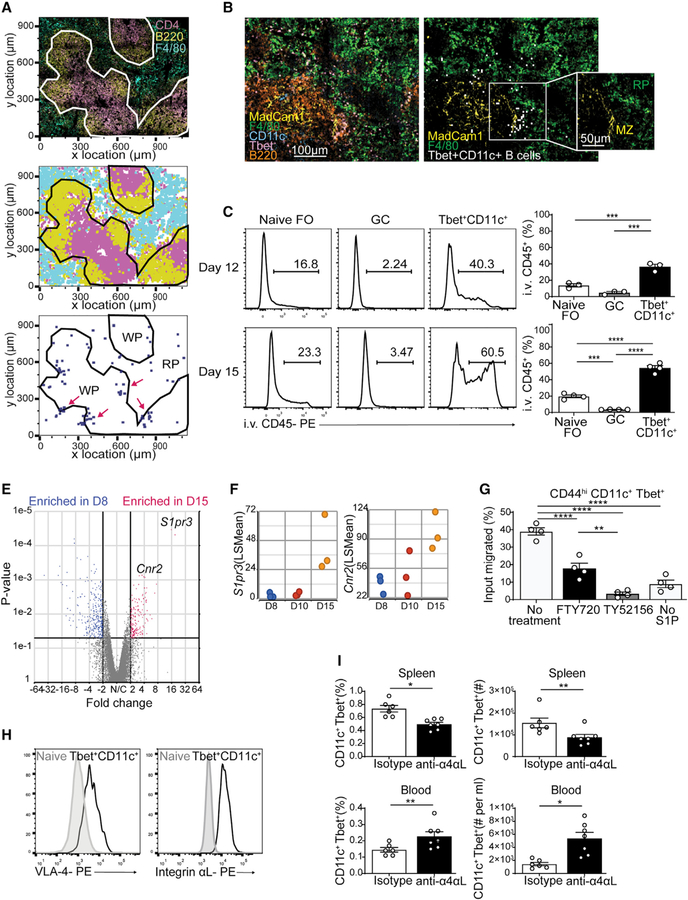
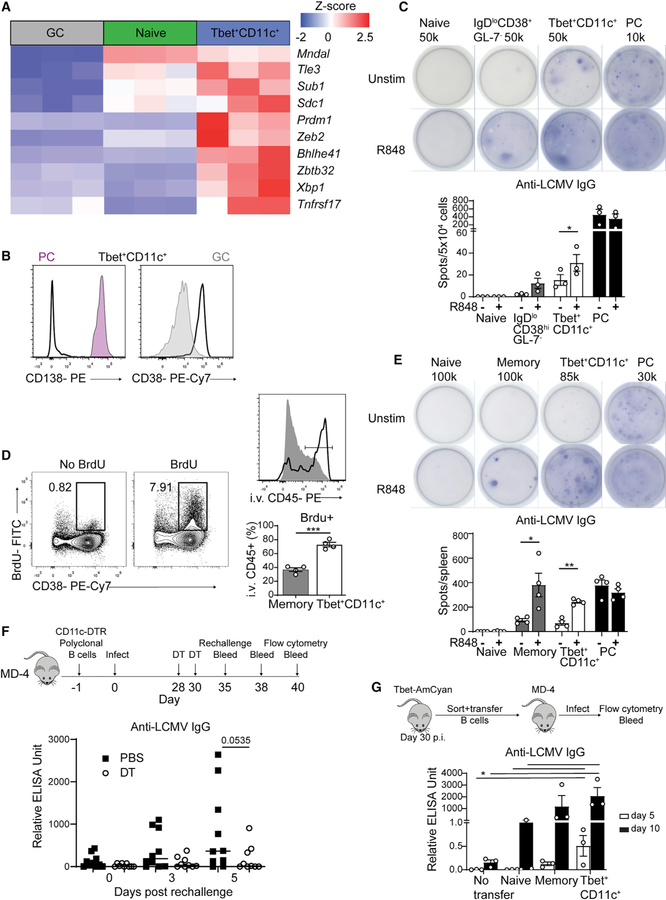
Tbet+CD11c+ B cells arise during type 1 pathogen challenge, aging, and autoimmunity in mice and humans. Here, we examined the developmental requirements of this B cell subset. In acute infection, T follicular helper (Tfh) cells, but not Th1 cells, drove Tbet+CD11c+ B cell generation through proximal delivery of help. Tbet+CD11c+ B cells developed prior to germinal center (GC) formation, exhibiting phenotypic and transcriptional profiles distinct from GC B cells. Fate tracking revealed that most Tbet+CD11c+ B cells developed independently of GC entry and cell-intrinsic Bcl6 expression. Tbet+CD11c+ and GC B cells exhibited minimal repertoire overlap, indicating distinct developmental pathways. As the infection resolved, Tbet+CD11c+ B cells localized to the marginal zone where splenic retention depended on integrins LFA-1 and VLA-4, forming a competitive memory subset that contributed to antibody production and secondary GC seeding upon rechallenge. Therefore, Tbet+CD11c+ B cells comprise a GC-independent memory subset capable of rapid and robust recall responses.
Keywords: Tbet(+)CD11c(+) B cells; Tfh cells; age-associated B cells; germinal center; humoral memory.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
"Are you gonna go my way?"-Decisions at the Tfh-B cell interface.Immunity. 2022 Mar 8;55(3):377-379. doi: 10.1016/j.immuni.2022.02.002. Epub 2022 Feb 17. Immunity. 2022. PMID: 35180377
References
-
- Austin JW, Buckner CM, Kardava L, Wang W, Zhang X, Melson VA, Swanson RG, Martins AJ, Zhou JQ, Hoehn KB, et al. (2019). Overexpression of T-bet in HIV infection is associated with accumulation of B cells outside germinal centers and poor affinity maturation. Sci. Transl. Med. 11, eaax0904. - PMC - PubMed
-
- Baddeley A, and Turner R (2005). spatstat: an R package for analyzing spatial point patterns. J. Stat. Softw. 12, 1–42.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
