

Refining the domain architecture model of the replication origin firing factor Treslin/TICRR
- PMID: 35091422
- PMCID: PMC8807876
- DOI: 10.26508/lsa.202101088
Refining the domain architecture model of the replication origin firing factor Treslin/TICRR
Abstract
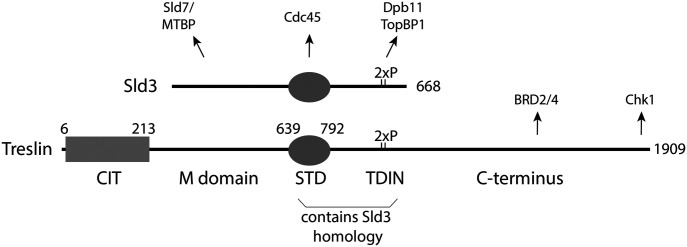
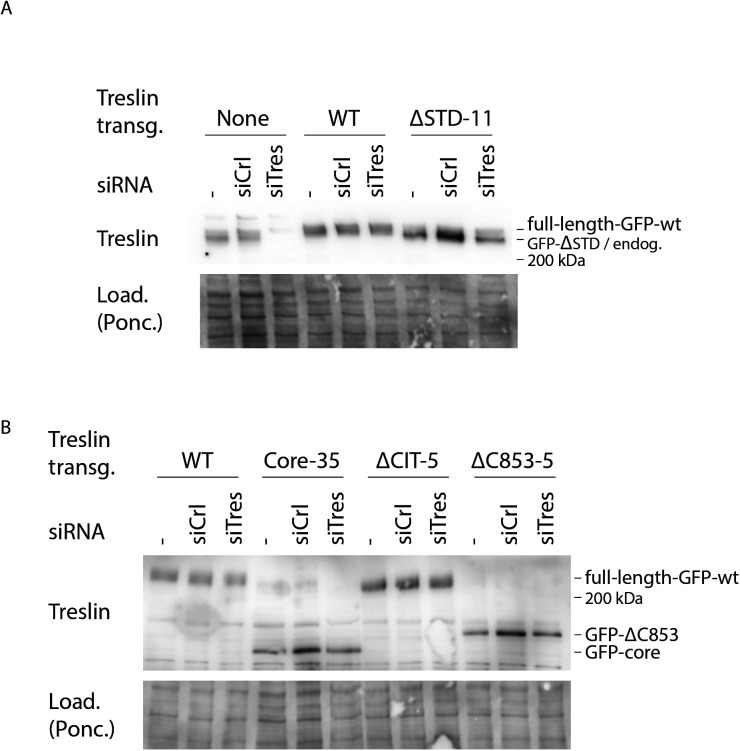
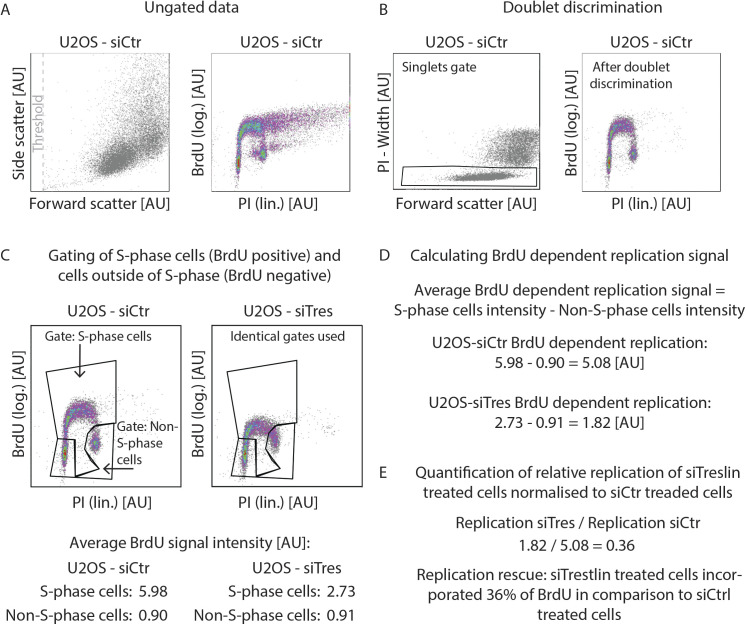
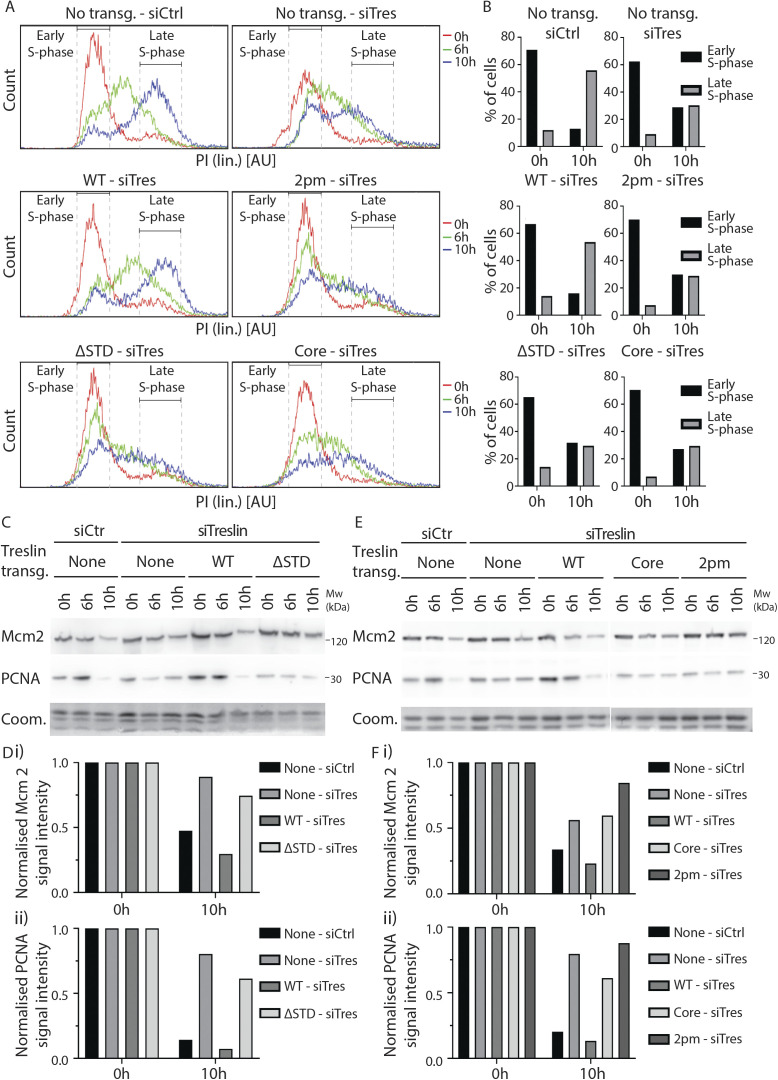
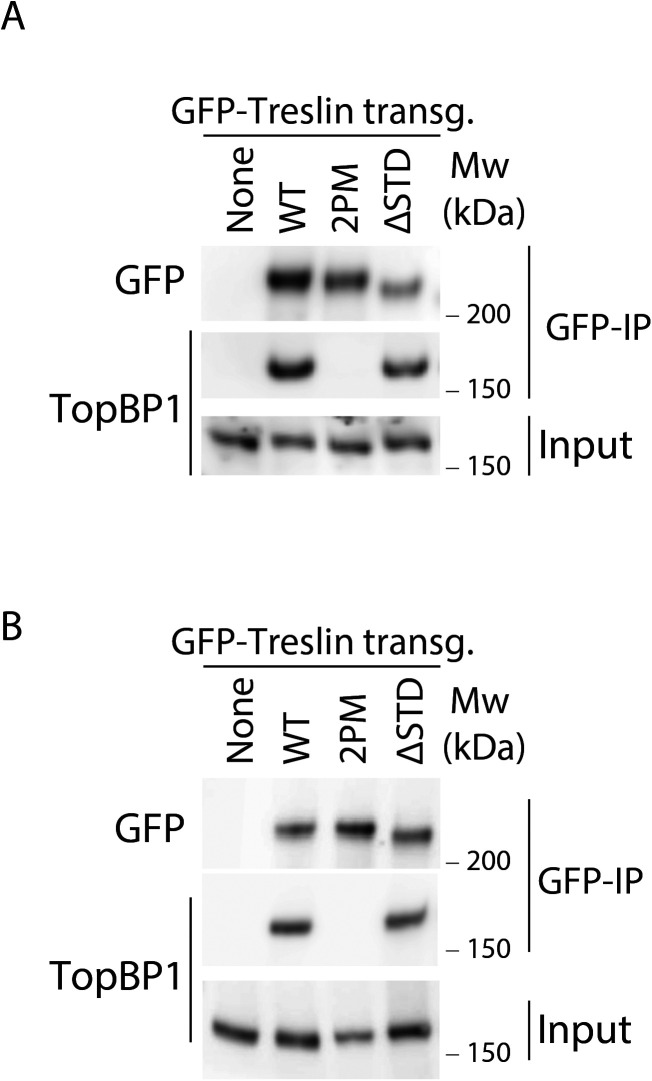
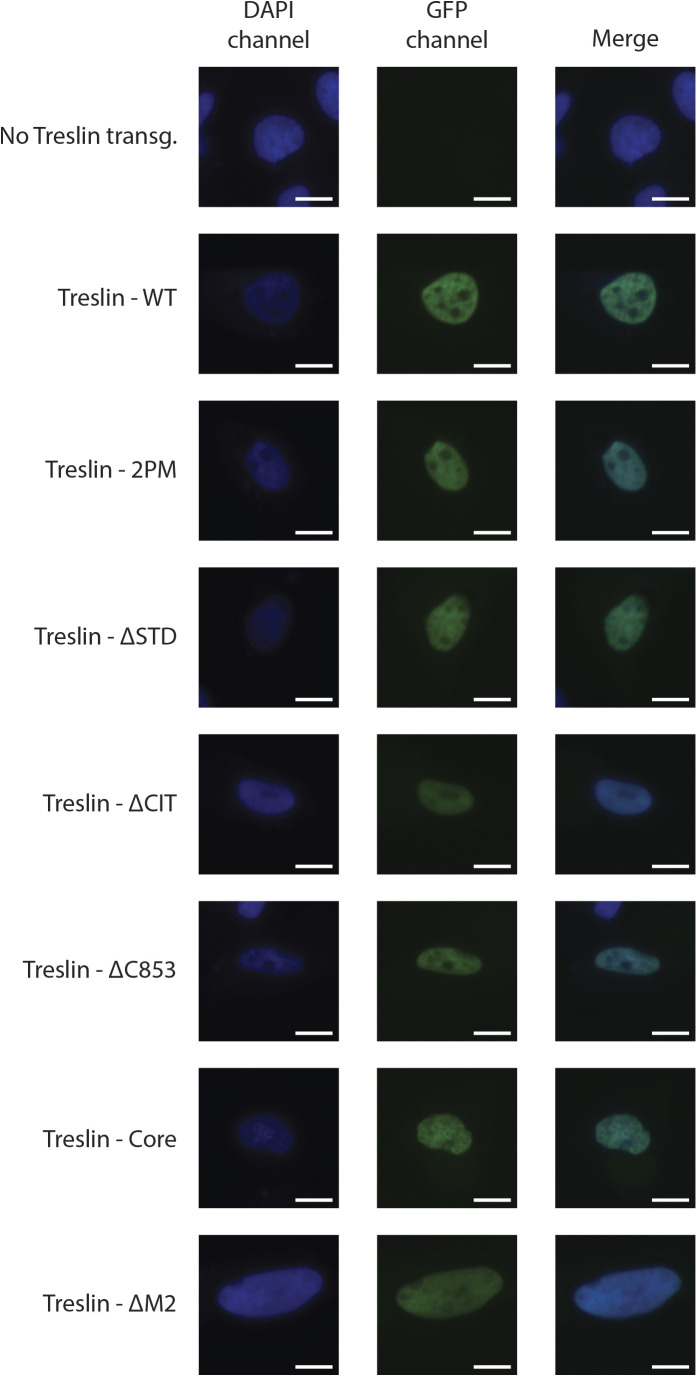
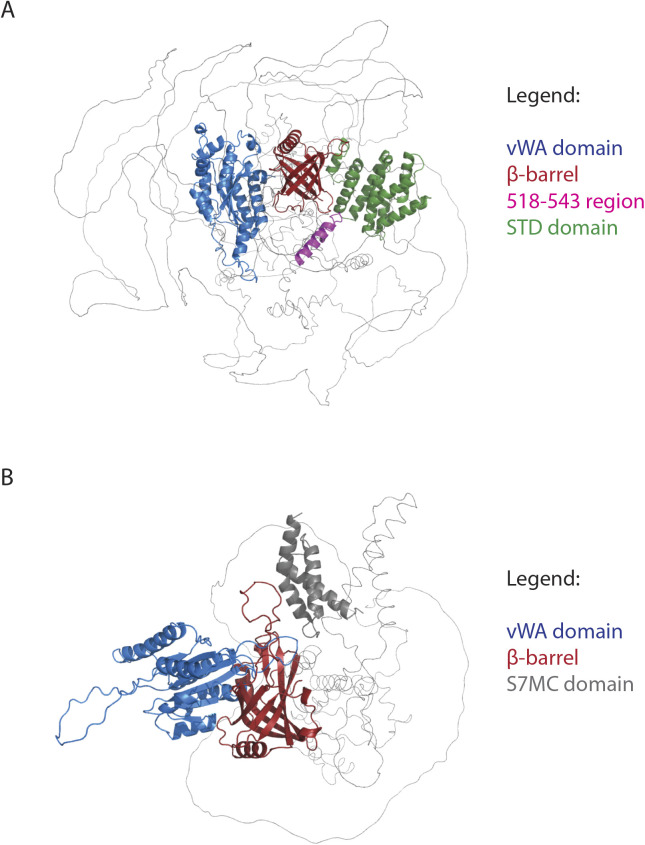
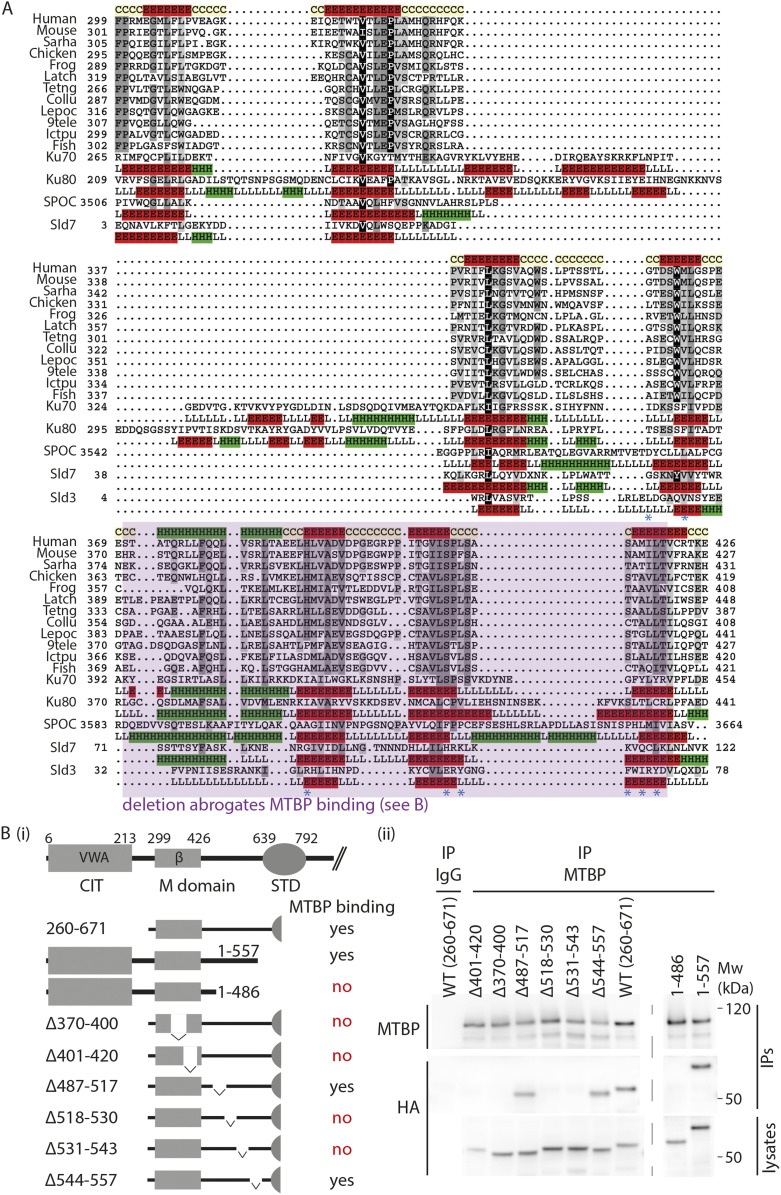
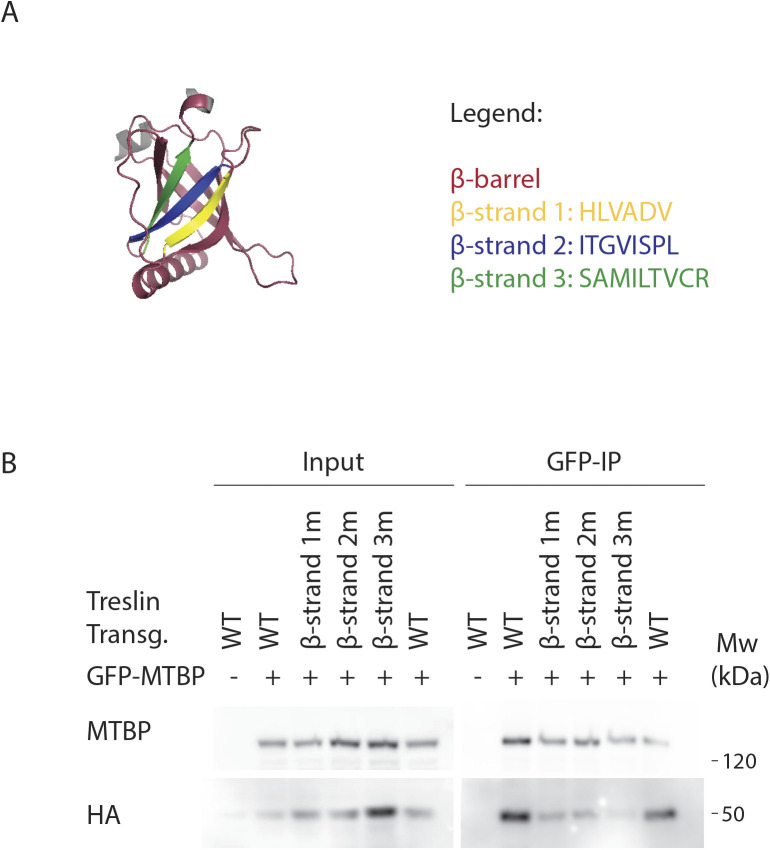
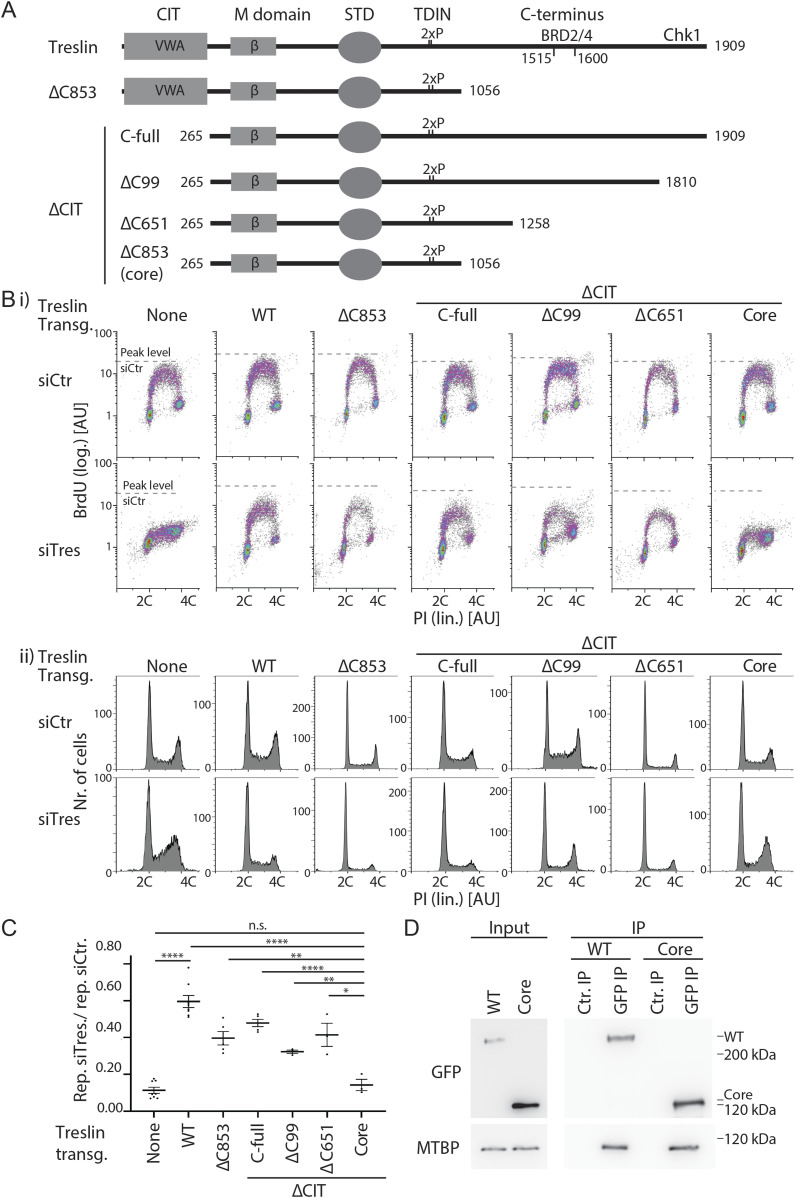
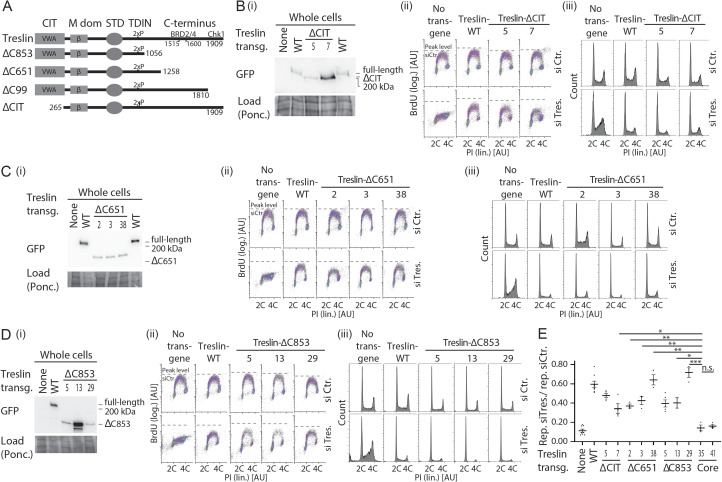
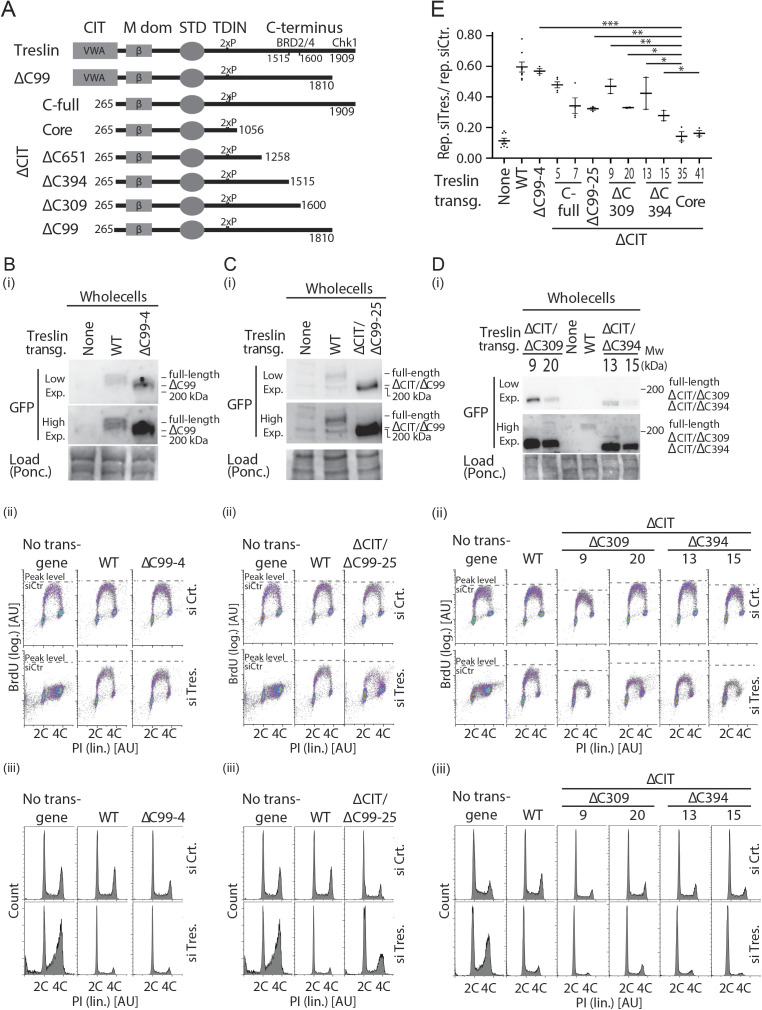
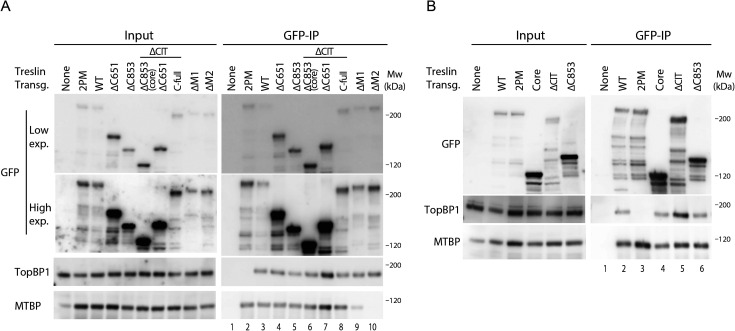
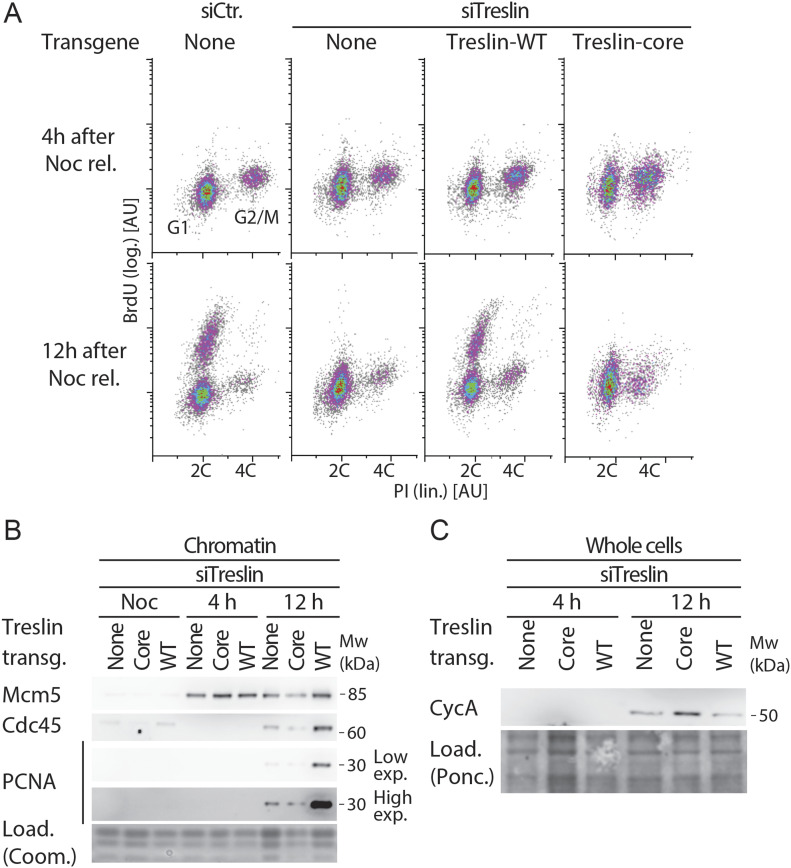
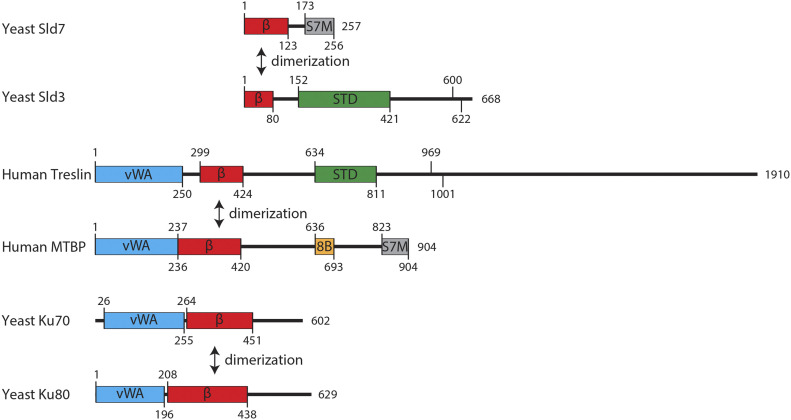
Faithful genome duplication requires appropriately controlled replication origin firing. The metazoan origin firing regulation hub Treslin/TICRR and its yeast orthologue Sld3 share the Sld3-Treslin domain and the adjacent TopBP1/Dpb11 interaction domain. We report a revised domain architecture model of Treslin/TICRR. Protein sequence analyses uncovered a conserved Ku70-homologous β-barrel fold in the Treslin/TICRR middle domain (M domain) and in Sld3. Thus, the Sld3-homologous Treslin/TICRR core comprises its three central domains, M domain, Sld3-Treslin domain, and TopBP1/Dpb11 interaction domain, flanked by non-conserved terminal domains, the CIT (conserved in Treslins) and the C terminus. The CIT includes a von Willebrand factor type A domain. Unexpectedly, MTBP, Treslin/TICRR, and Ku70/80 share the same N-terminal domain architecture, von Willebrand factor type A and Ku70-like β-barrels, suggesting a common ancestry. Binding experiments using mutants and the Sld3-Sld7 dimer structure suggest that the Treslin/Sld3 and MTBP/Sld7 β-barrels engage in homotypic interactions, reminiscent of Ku70-Ku80 dimerization. Cells expressing Treslin/TICRR domain mutants indicate that all Sld3-core domains and the non-conserved terminal domains fulfil important functions during origin firing in human cells. Thus, metazoa-specific and widely conserved molecular processes cooperate during metazoan origin firing.
© 2022 Ferreira et al.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures




References
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous