

The Hippo pathway effectors YAP and TAZ interact with EGF-like signaling to regulate expansion-related events in bovine cumulus cells in vitro
- PMID: 35091965
- PMCID: PMC8956774
- DOI: 10.1007/s10815-021-02384-x
The Hippo pathway effectors YAP and TAZ interact with EGF-like signaling to regulate expansion-related events in bovine cumulus cells in vitro
Abstract
Purpose: To determine if the inhibition of the interaction between the Hippo effector YAP or its transcriptional co-activator TAZ with the TEAD family of transcription factors is critical for the cumulus expansion-related events induced by the EGF network in cumulus-oocyte complexes (COCs).
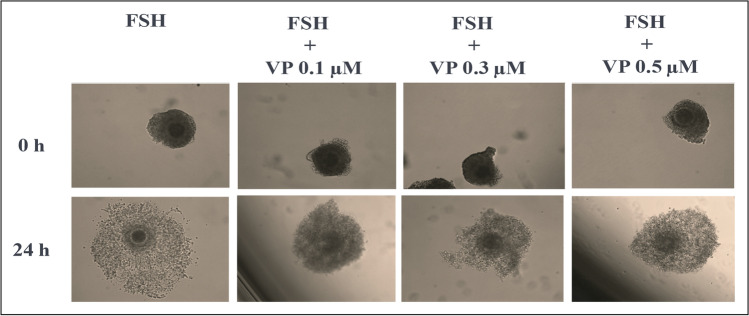
Methods: We performed a series of experiments using immature bovine COCs subjected to an IVM protocol for up 24 h in which cumulus expansion was stimulated with EGF recombinant protein or FSH.
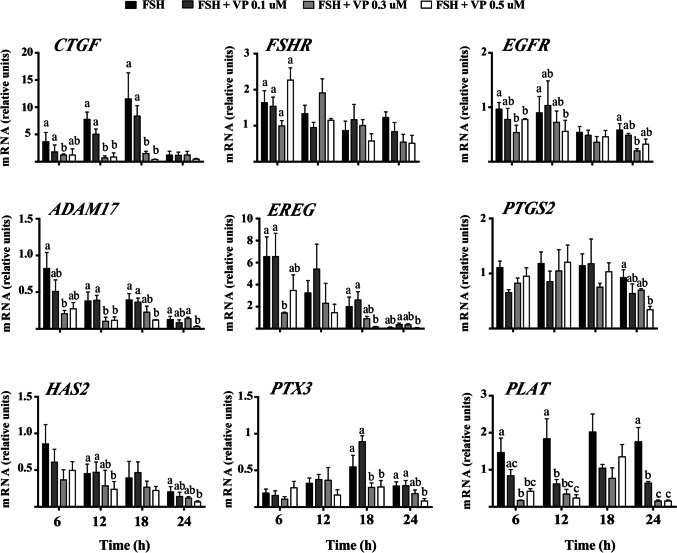
Results: The main results indicated that EGFR activity stimulation in bovine cumulus cells (CC) increases mRNA levels encoding the classic YAP/TAZ-TEAD target gene CTGF. To determine if important genes for cumulus expansion are transcriptional targets of YAP/TAZ-TEAD interaction in CC, COCs were then subjected to IVM in the presence of FSH with or without distinct concentrations of Verteporfin (VP; a small molecule inhibitor that interferes with YAP/TAZ binding to TEADs). COCs were then collected at 6, 12, 18, and 24 h for total RNA extraction and RT-qPCR analyses. This experiment indicated that VP inhibits in a time- and concentration-dependent manner distinct cumulus expansion and oocyte maturation-related genes, by regulating EGFR and CTGF expression in CC.
Conclusions: Taken together, the results presented herein represent considerable insight into the functional relevance of a completely novel signaling pathway underlying cumulus expansion and oocyte maturation in monovulatory species. YAP/TAZ or CTGF may represent potential targets to improve the efficiency of IVM systems, not only for monovulatory species of agricultural importance as the cow, but for human embryo production.
Keywords: CTGF; Cow; EGF receptor; Oocyte in vitro maturation; Ovary; Verteporfin.
© 2021. The Author(s), under exclusive licence to Springer Science+Business Media, LLC, part of Springer Nature.
Conflict of interest statement
The authors declare no competing interests.
Figures



Similar articles
-
FSH Regulates YAP-TEAD Transcriptional Activity in Bovine Granulosa Cells to Allow the Future Dominant Follicle to Exert Its Augmented Estrogenic Capacity.Int J Mol Sci. 2022 Nov 16;23(22):14160. doi: 10.3390/ijms232214160. Int J Mol Sci. 2022. PMID: 36430640 Free PMC article.
-
Oocyte secreted factors control genes regulating FSH signaling and the maturation cascade in cumulus cells: the oocyte is not in a hurry.J Assist Reprod Genet. 2023 Aug;40(8):1961-1971. doi: 10.1007/s10815-023-02822-y. Epub 2023 May 19. J Assist Reprod Genet. 2023. PMID: 37204638 Free PMC article.
-
NPR3 is regulated by gonadotropins and modulates bovine cumulus cell expansion.Reproduction. 2024 Dec 9;169(1):e240187. doi: 10.1530/REP-24-0187. Print 2025 Jan 1. Reproduction. 2024. PMID: 39441769
-
Regulation of cumulus expansion and hyaluronan synthesis in porcine oocyte-cumulus complexes during in vitro maturation.Endocr Regul. 2012 Oct;46(4):225-35. doi: 10.4149/endo_2012_04_225. Endocr Regul. 2012. PMID: 23127506 Review.
-
Emerging roles for the YAP/TAZ transcriptional regulators in brain tumour pathology and targeting options.Neuropathol Appl Neurobiol. 2022 Feb;48(2):e12762. doi: 10.1111/nan.12762. Epub 2021 Aug 26. Neuropathol Appl Neurobiol. 2022. PMID: 34409639 Review.
Cited by
-
The Effect of Stimulation Protocols (GnRH Agonist vs. Antagonist) on the Activity of mTOR and Hippo Pathways of Ovarian Granulosa Cells and Its Potential Correlation with the Outcomes of In Vitro Fertilization: A Hypothesis.J Clin Med. 2022 Oct 18;11(20):6131. doi: 10.3390/jcm11206131. J Clin Med. 2022. PMID: 36294452 Free PMC article.
-
Is the Hippo Pathway Effector Yes-Associated Protein a Potential Key Player of Dairy Cattle Cystic Ovarian Disease Pathogenesis?Animals (Basel). 2023 Sep 8;13(18):2851. doi: 10.3390/ani13182851. Animals (Basel). 2023. PMID: 37760251 Free PMC article.
-
FSH Regulates YAP-TEAD Transcriptional Activity in Bovine Granulosa Cells to Allow the Future Dominant Follicle to Exert Its Augmented Estrogenic Capacity.Int J Mol Sci. 2022 Nov 16;23(22):14160. doi: 10.3390/ijms232214160. Int J Mol Sci. 2022. PMID: 36430640 Free PMC article.
-
Sustained YAP1 signaling alters the fates of lutein and Müllerian mesenchymal cells in mice.Reproduction. 2025 Jul 24;170(2):e240450. doi: 10.1530/REP-24-0450. Print 2025 Aug 1. Reproduction. 2025. PMID: 40658769 Free PMC article.
References
-
- Bromer JG, Cetinkaya MB, Arici A: Pretreatments before the induction of ovulation in assisted reproduction technologies: evidence-based medicine in 2007. Ann N Y Acad Sci 2008, 1127 31–40 10.1196/annals.1434.004 - PubMed
-
- Shimada M, Hernandez-Gonzalez I, Gonzalez-Robayna I, Richards JS. Paracrine and autocrine regulation of epidermal growth factor-like factors in cumulus oocyte complexes and granulosa cells: key roles for prostaglandin synthase 2 and progesterone receptor. Mol Endocrinol. 2006;20(6):1352–1365. doi: 10.1210/me.2005-0504. - DOI - PubMed
MeSH terms
Substances
Grants and funding
- Discovery Grant RGPIN-2018-06470/natural sciences and engineering research council of canada
- Edital 06/2019/fundação de amparo à pesquisa do estado do rio grande do sul
- 19/2551-0002275-1/fundação de amparo à pesquisa do estado do rio grande do sul
- 16/2551-0000494-3/conselho nacional de desenvolvimento científico e tecnológico
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
