

Domestication Impacts the Wheat-Associated Microbiota and the Rhizosphere Colonization by Seed- and Soil-Originated Microbiomes, Across Different Fields
- PMID: 35095978
- PMCID: PMC8789879
- DOI: 10.3389/fpls.2021.806915
Domestication Impacts the Wheat-Associated Microbiota and the Rhizosphere Colonization by Seed- and Soil-Originated Microbiomes, Across Different Fields
Abstract
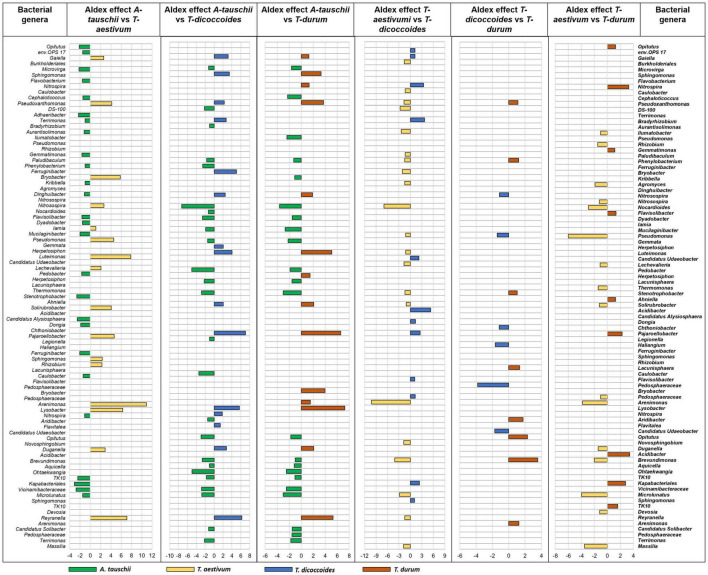
The seed-transmitted microorganisms and the microbiome of the soil in which the plant grows are major drivers of the rhizosphere microbiome, a crucial component of the plant holobiont. The seed-borne microbiome can be even coevolved with the host plant as a result of adaptation and vertical transmission over generations. The reduced genome diversity and crossing events during domestication might have influenced plant traits that are important for root colonization by seed-borne microbes and also rhizosphere recruitment of microbes from the bulk soil. However, the impact of the breeding on seed-transmitted microbiome composition and the plant ability of microbiome selection from the soil remain unknown. Here, we analyzed both endorhiza and rhizosphere microbiome of two couples of genetically related wild and cultivated wheat species (Aegilops tauschii/Triticum aestivum and T. dicoccoides/T. durum) grown in three locations, using 16S rRNA gene and ITS2 metabarcoding, to assess the relative contribution of seed-borne and soil-derived microbes to the assemblage of the rhizosphere microbiome. We found that more bacterial and fungal ASVs are transmitted from seed to the endosphere of all species compared with the rhizosphere, and these transmitted ASVs were species-specific regardless of location. Only in one location, more microbial seed transmission occurred also in the rhizosphere of A. tauschii compared with other species. Concerning soil-derived microbiome, the most distinct microbial genera occurred in the rhizosphere of A. tauschii compared with other species in all locations. The rhizosphere of genetically connected wheat species was enriched with similar taxa, differently between locations. Our results demonstrate that host plant criteria for soil bank's and seed-originated microbiome recruitment depend on both plants' genotype and availability of microorganisms in a particular environment. This study also provides indications of coevolution between the host plant and its associated microbiome resulting from the vertical transmission of seed-originated taxa.
Keywords: bulk soil; coevolution; crop domestication; rhizosphere; seed microbiome.
Copyright © 2022 Abdullaeva, Ratering, Ambika Manirajan, Rosado-Porto, Schnell and Cardinale.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







References
-
- Aitchison J. (1986). The Statistical Analysis of Compositional Data. London: Chapman and Hall.
-
- Aitchison J., Greenacre M. (2002). Biplots of compositional data. J. R. Stat. Soc. Ser. C Appl. Stat. 51 375–392.
LinkOut - more resources
Full Text Sources