

Transcription factor regulation of eQTL activity across individuals and tissues
- PMID: 35100260
- PMCID: PMC8830792
- DOI: 10.1371/journal.pgen.1009719
Transcription factor regulation of eQTL activity across individuals and tissues
Abstract
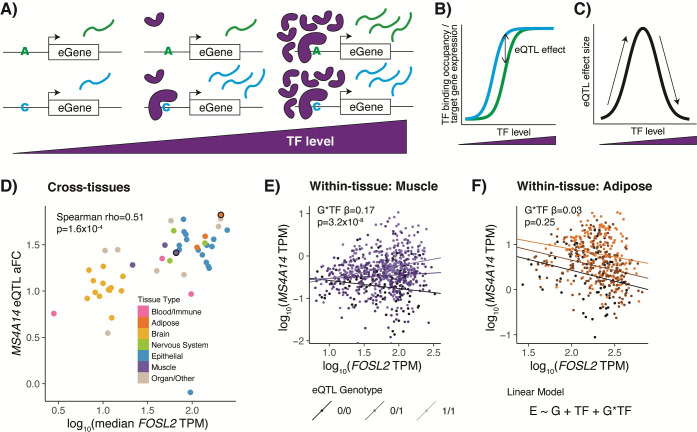
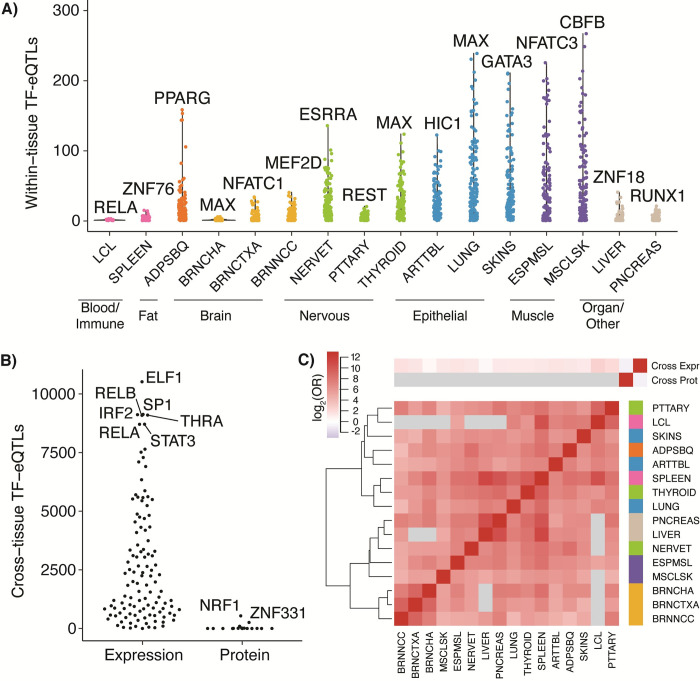
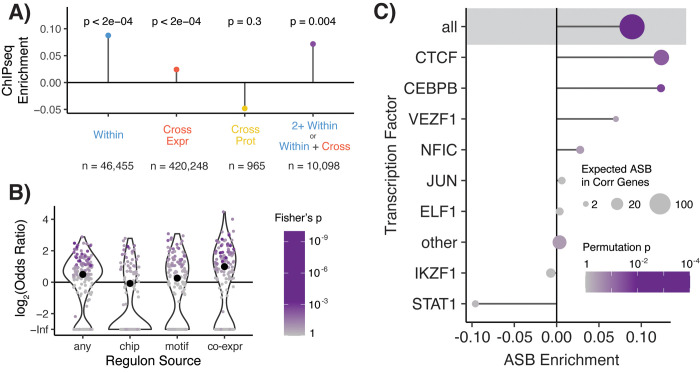
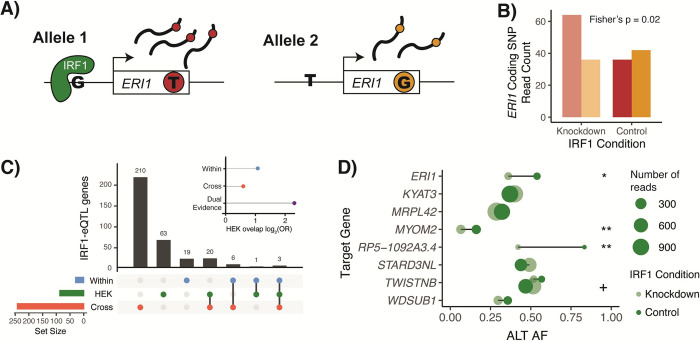
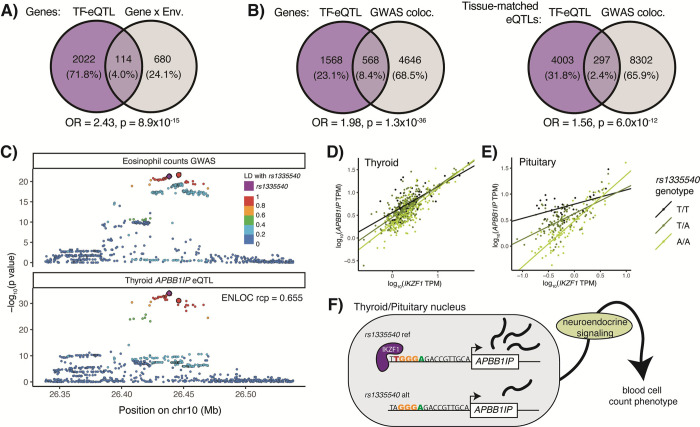
Tens of thousands of genetic variants associated with gene expression (cis-eQTLs) have been discovered in the human population. These eQTLs are active in various tissues and contexts, but the molecular mechanisms of eQTL variability are poorly understood, hindering our understanding of genetic regulation across biological contexts. Since many eQTLs are believed to act by altering transcription factor (TF) binding affinity, we hypothesized that analyzing eQTL effect size as a function of TF level may allow discovery of mechanisms of eQTL variability. Using GTEx Consortium eQTL data from 49 tissues, we analyzed the interaction between eQTL effect size and TF level across tissues and across individuals within specific tissues and generated a list of 10,098 TF-eQTL interactions across 2,136 genes that are supported by at least two lines of evidence. These TF-eQTLs were enriched for various TF binding measures, supporting with orthogonal evidence that these eQTLs are regulated by the implicated TFs. We also found that our TF-eQTLs tend to overlap genes with gene-by-environment regulatory effects and to colocalize with GWAS loci, implying that our approach can help to elucidate mechanisms of context-specificity and trait associations. Finally, we highlight an interesting example of IKZF1 TF regulation of an APBB1IP gene eQTL that colocalizes with a GWAS signal for blood cell traits. Together, our findings provide candidate TF mechanisms for a large number of eQTLs and offer a generalizable approach for researchers to discover TF regulators of genetic variant effects in additional QTL datasets.
Conflict of interest statement
I have read the journal’s policy and the authors of this manuscript have the following competing interests: EDF is currently employed by Patch Biosciences. TL advises Variant Bio, Goldfinch Bio, GlaxoSmithKline and has equity in Variant Bio. FA is an inventor on a patent application related to TensorQTL.
Figures
Similar articles
-
reg-eQTL: Integrating transcription factor effects to unveil regulatory variants.Am J Hum Genet. 2025 Mar 6;112(3):659-674. doi: 10.1016/j.ajhg.2025.01.015. Epub 2025 Feb 7. Am J Hum Genet. 2025. PMID: 39922197 Free PMC article.
-
Tissue specific regulation of transcription in endometrium and association with disease.Hum Reprod. 2020 Feb 29;35(2):377-393. doi: 10.1093/humrep/dez279. Hum Reprod. 2020. PMID: 32103259 Free PMC article.
-
Characterizing the Relation Between Expression QTLs and Complex Traits: Exploring the Role of Tissue Specificity.Behav Genet. 2018 Sep;48(5):374-385. doi: 10.1007/s10519-018-9914-2. Epub 2018 Jul 20. Behav Genet. 2018. PMID: 30030655 Free PMC article.
-
Expression Quantitative Trait Loci Information Improves Predictive Modeling of Disease Relevance of Non-Coding Genetic Variation.PLoS One. 2015 Oct 16;10(10):e0140758. doi: 10.1371/journal.pone.0140758. eCollection 2015. PLoS One. 2015. PMID: 26474488 Free PMC article. Review.
-
QTL Analysis Beyond eQTLs.Methods Mol Biol. 2020;2082:201-210. doi: 10.1007/978-1-0716-0026-9_14. Methods Mol Biol. 2020. PMID: 31849017 Review.
Cited by
-
SNP rs615552 and lncRNA CDKN2B-AS1 influence brain cancer pathogenesis through multi-omic mechanisms.Sci Rep. 2025 Jul 28;15(1):27490. doi: 10.1038/s41598-025-10360-z. Sci Rep. 2025. PMID: 40721593 Free PMC article.
-
Microbiome-associated human genetic variants impact phenome-wide disease risk.Proc Natl Acad Sci U S A. 2022 Jun 28;119(26):e2200551119. doi: 10.1073/pnas.2200551119. Epub 2022 Jun 24. Proc Natl Acad Sci U S A. 2022. PMID: 35749358 Free PMC article.
-
Functional characterization of human genomic variation linked to polygenic diseases.Trends Genet. 2023 Jun;39(6):462-490. doi: 10.1016/j.tig.2023.02.014. Epub 2023 Mar 28. Trends Genet. 2023. PMID: 36997428 Free PMC article. Review.
-
Single-cell eQTL analysis identifies genetic variation underlying metabolic dysfunction-associated steatohepatitis.Nat Genet. 2025 Jul;57(7):1638-1648. doi: 10.1038/s41588-025-02237-8. Epub 2025 Jun 25. Nat Genet. 2025. PMID: 40562914
-
Role of expression quantitative trait loci (eQTL) in understanding genetic mechanisms underlying common complex diseases.Mol Cells. 2025 Sep;48(9):100256. doi: 10.1016/j.mocell.2025.100256. Epub 2025 Jul 18. Mol Cells. 2025. PMID: 40684919 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
