

Chromosome-level genome assembly of the fully mycoheterotrophic orchid Gastrodia elata
- PMID: 35100375
- PMCID: PMC8896018
- DOI: 10.1093/g3journal/jkab433
Chromosome-level genome assembly of the fully mycoheterotrophic orchid Gastrodia elata
Abstract
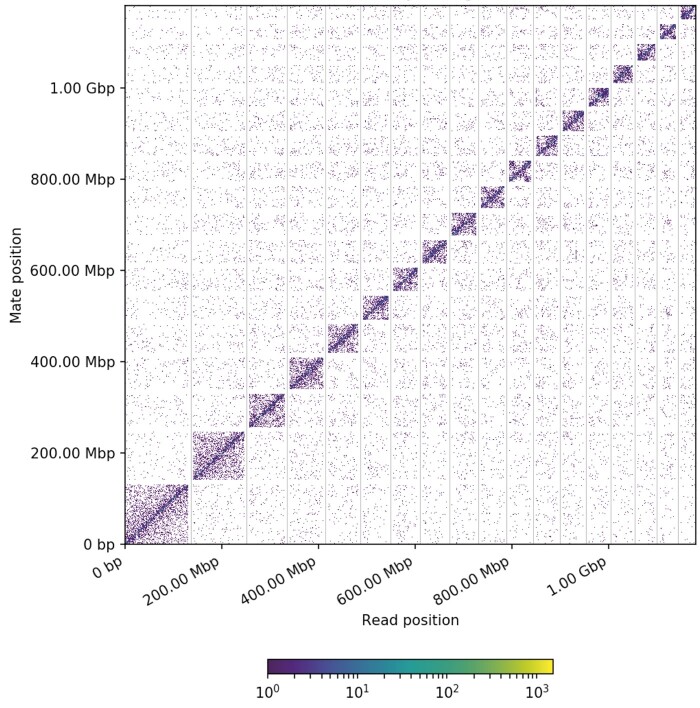
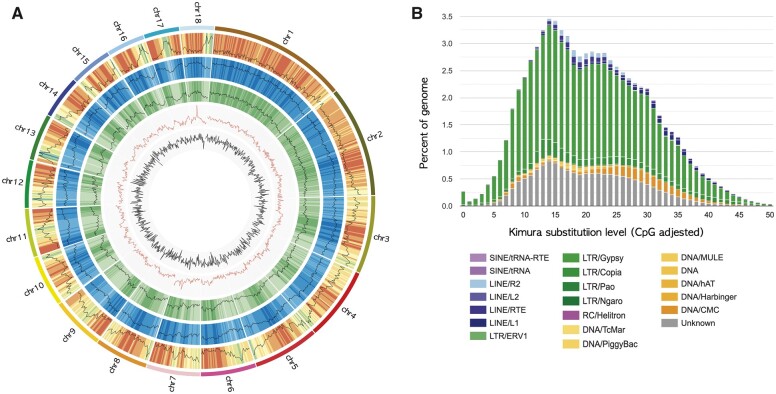
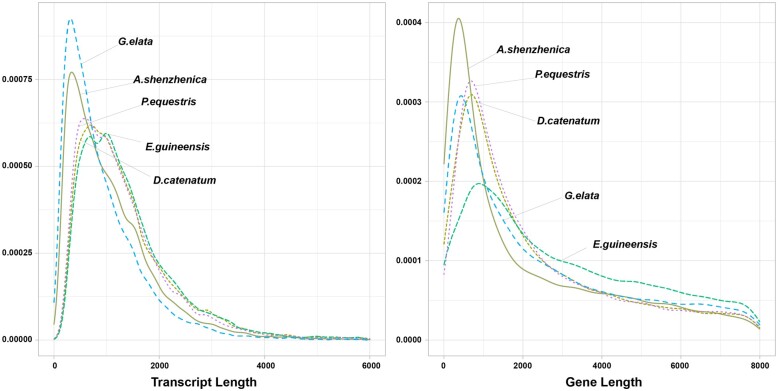
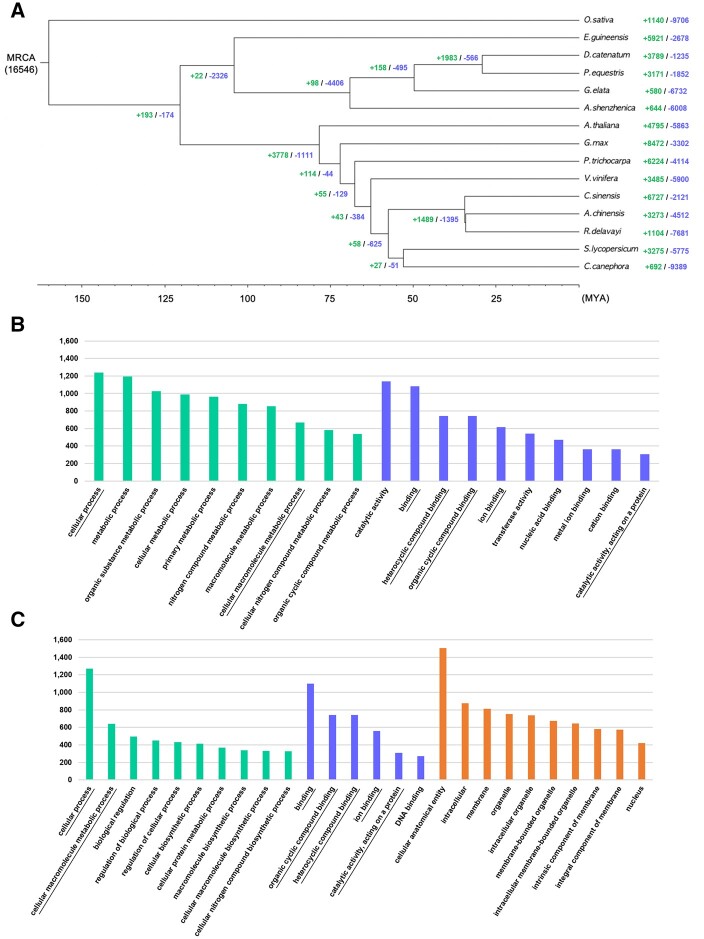
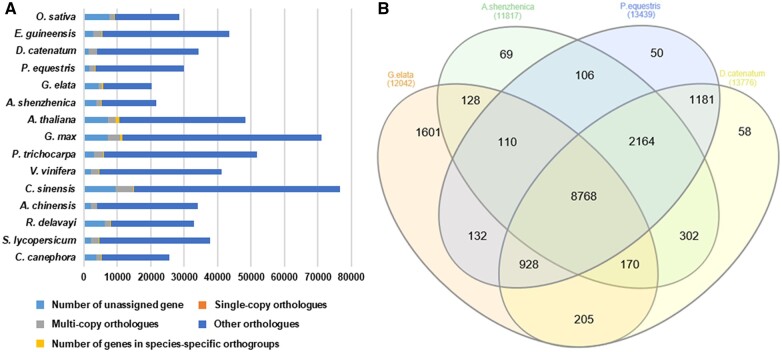
Gastrodia elata, an obligate mycoheterotrophic orchid, requires complete carbon and mineral nutrient supplementation from mycorrhizal fungi during its entire life cycle. Although full mycoheterotrophy occurs most often in the Orchidaceae family, no chromosome-level reference genome from this group has been assembled to date. Here, we report a high-quality chromosome-level genome assembly of G. elata, using Illumina and PacBio sequencing methods with Hi-C technique. The assembled genome size was found to be 1045 Mb, with an N50 of 50.6 Mb and 488 scaffolds. A total of 935 complete (64.9%) matches to the 1440 embryophyte Benchmarking Universal Single-Copy Orthologs were identified in this genome assembly. Hi-C scaffolding of the assembled genome resulted in 18 pseudochromosomes, 1008 Mb in size and containing 96.5% of the scaffolds. A total of 18,844 protein-coding sequences (CDSs) were predicted in the G. elata genome, of which 15,619 CDSs (82.89%) were functionally annotated. In addition, 74.92% of the assembled genome was found to be composed of transposable elements. Phylogenetic analysis indicated a significant contraction of genes involved in various biosynthetic processes and cellular components and an expansion of genes for novel metabolic processes and mycorrhizal association. This result suggests an evolutionary adaptation of G. elata to a mycoheterotrophic lifestyle. In summary, the genomic resources generated in this study will provide a valuable reference genome for investigating the molecular mechanisms of G. elata biological functions. Furthermore, the complete G. elata genome will greatly improve our understanding of the genetics of Orchidaceae and its mycoheterotrophic evolution.
Keywords: Gastrodia elata; Orchidaceae; genome assembly; mycoheterotrophic; pseudochromosome.
© The Author(s) 2022. Published by Oxford University Press on behalf of Genetics Society of America.
Figures

References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous