

Force-Induced Changes of PilY1 Drive Surface Sensing by Pseudomonas aeruginosa
- PMID: 35100866
- PMCID: PMC8806160
- DOI: 10.1128/mbio.03754-21
Force-Induced Changes of PilY1 Drive Surface Sensing by Pseudomonas aeruginosa
Abstract
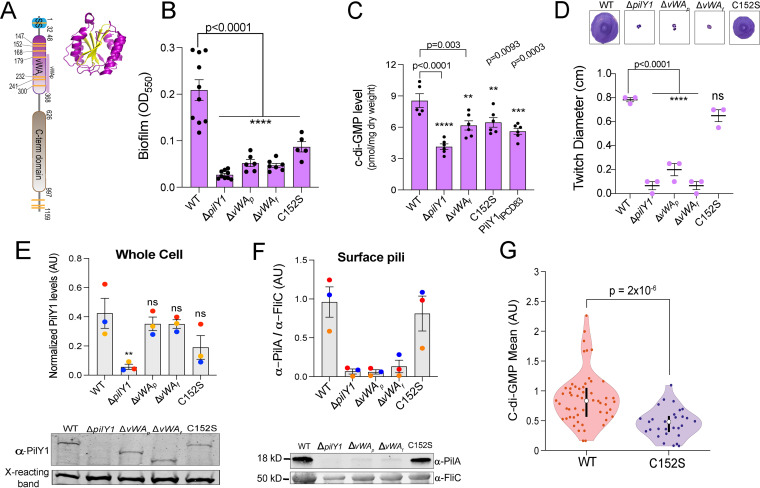
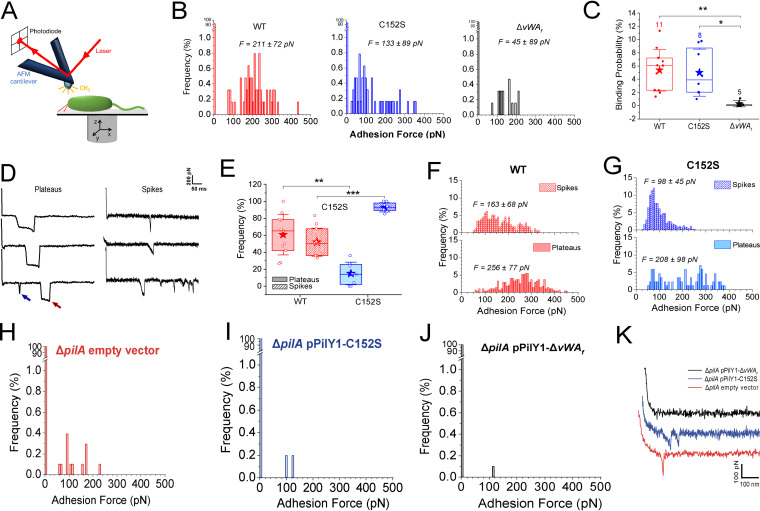
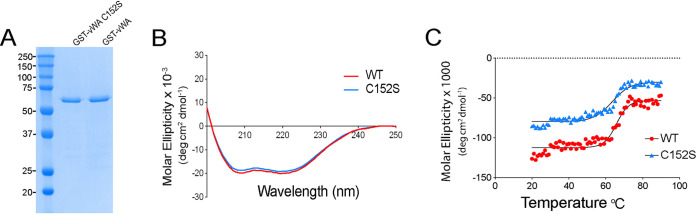
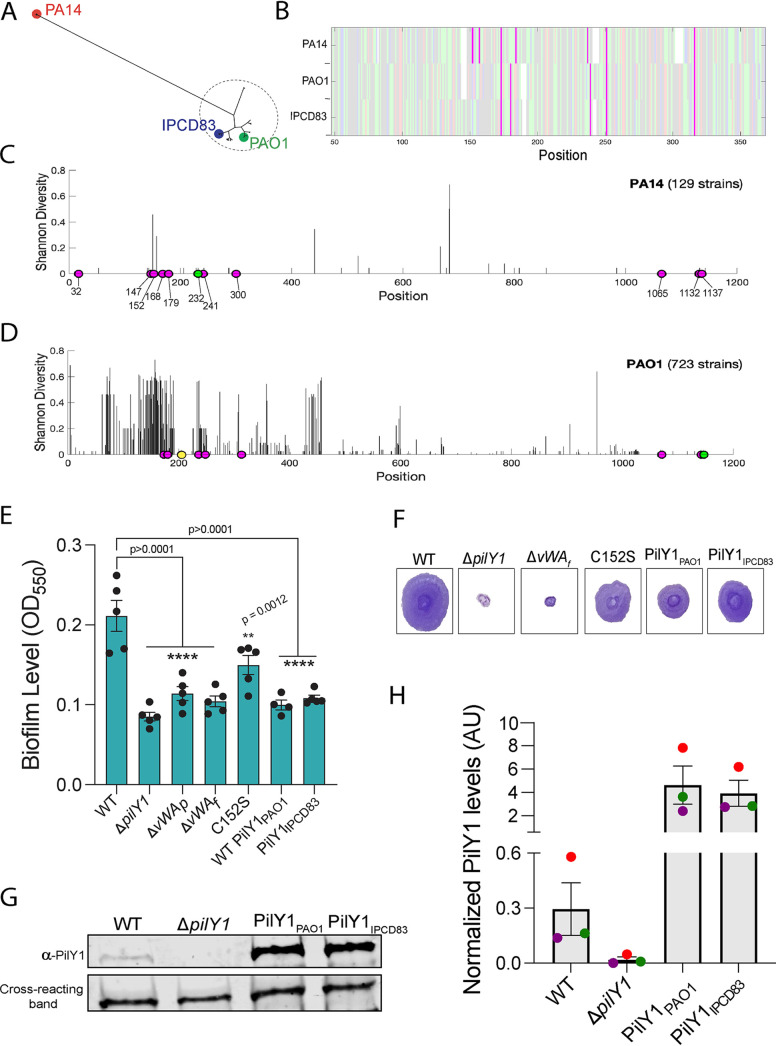
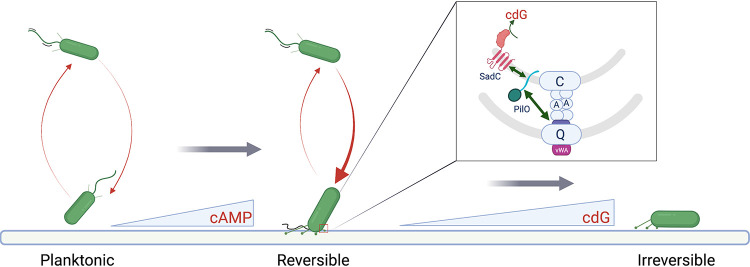
During biofilm formation, the opportunistic pathogen Pseudomonas aeruginosa uses its type IV pili (TFP) to sense a surface, eliciting increased second-messenger production and regulating target pathways required to adapt to a surface lifestyle. The mechanisms whereby TFP detect surface contact are still poorly understood, although mechanosensing is often invoked, with few data supporting this claim. Using a combination of molecular genetics and single-cell analysis, with biophysical, biochemical, and genomics techniques, we show that force-induced changes mediated by the von Willebrand A (vWA) domain-containing, TFP tip-associated protein PilY1 are required for surface sensing. Atomic force microscopy shows that TFP/PilY1 can undergo force-induced, sustained conformational changes akin to those observed for mechanosensitive proteins like titin. We show that mutation of a single cysteine residue in the vWA domain of PilY1 results in modestly lower surface adhesion forces, reduced sustained conformational changes, and increased nanospring-like properties, as well as reduced c-di-GMP signaling and biofilm formation. Mutating this cysteine has allowed us to genetically separate a role for TFP in twitching motility from surface-sensing signaling. The conservation of this Cys residue in all P. aeruginosa PA14 strains and its absence in the ∼720 sequenced strains of P. aeruginosa PAO1 may contribute to explaining the observed differences in surface colonization strategies observed for PA14 versus PAO1. IMPORTANCE Most bacteria live on abiotic and biotic surfaces in surface-attached communities known as biofilms. Surface sensing and increased levels of the second-messenger molecule c-di-GMP are crucial to the transition from planktonic to biofilm growth. The mechanism(s) underlying TFP-mediated surface detection that triggers this c-di-GMP signaling cascade is unclear. Here, we provide key insight into this question; we show that the eukaryote-like vWA domain of the TFP tip-associated protein PilY1 responds to mechanical force, which in turn drives the production of a key second messenger needed to regulate surface behaviors. Our studies highlight a potential mechanism that may account for differing surface colonization strategies.
Keywords: PilY1; c-di-GMP; force; surface sensing; type 4 pili; von Willebrand A domain.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Rodesney CA, Roman B, Dhamani N, Cooley BJ, Katira P, Touhami A, Gordon VD. 2017. Mechanosensing of shear by Pseudomonas aeruginosa leads to increased levels of the cyclic-di-GMP signal initiating biofilm development. Proc Natl Acad Sci U S A 114:5906–5911. doi:10.1073/pnas.1703255114. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
