

Novel insights into PORCN mutations, associated phenotypes and pathophysiological aspects
- PMID: 35101074
- PMCID: PMC8802438
- DOI: 10.1186/s13023-021-02068-w
Novel insights into PORCN mutations, associated phenotypes and pathophysiological aspects
Abstract
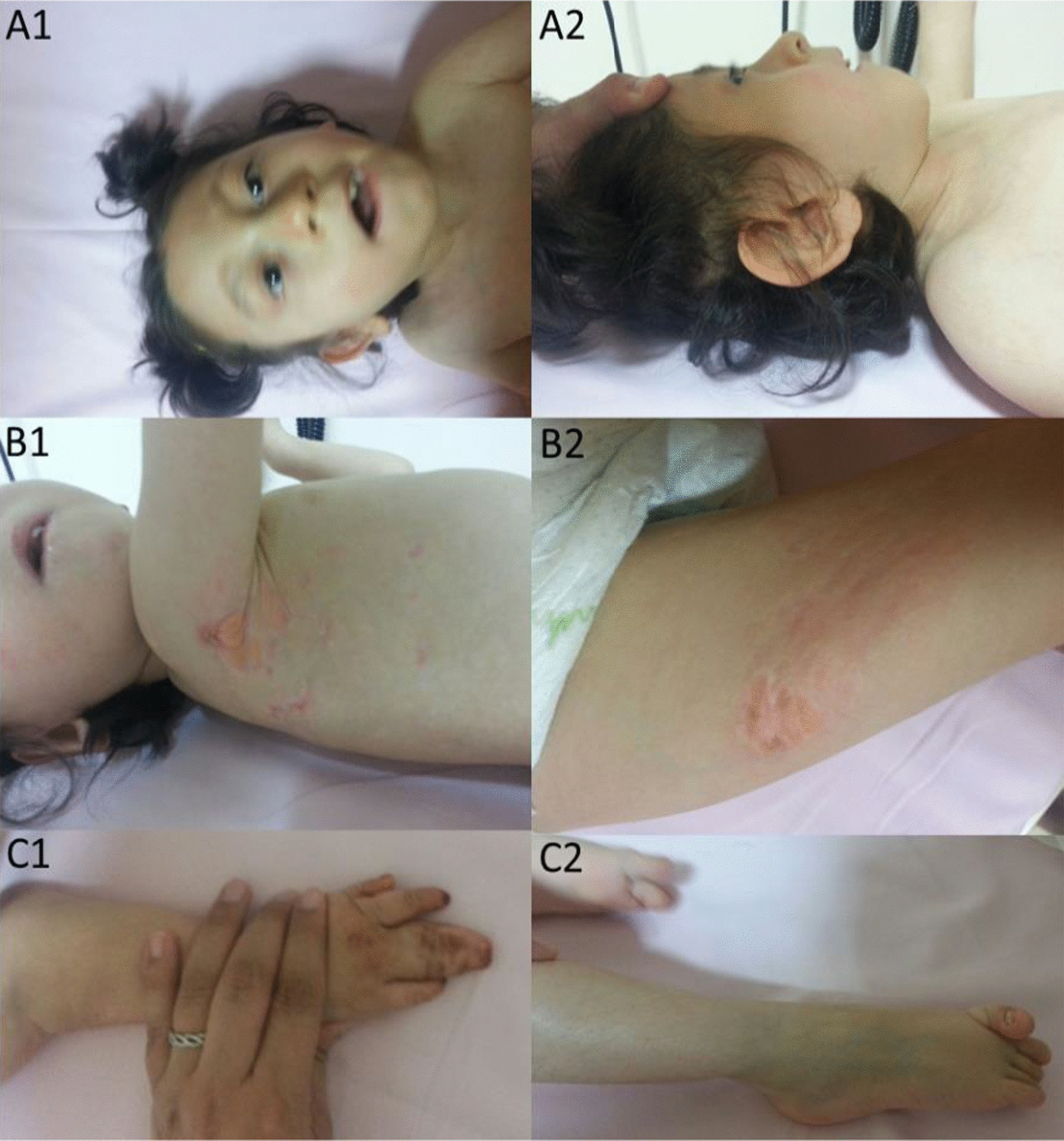
Background: Goltz syndrome (GS) is a X-linked disorder defined by defects of mesodermal- and ectodermal-derived structures and caused by PORCN mutations. Features include striated skin-pigmentation, ocular and skeletal malformations and supernumerary or hypoplastic nipples. Generally, GS is associated with in utero lethality in males and most of the reported male patients show mosaicism (only three non-mosaic surviving males have been described so far). Also, precise descriptions of neurological deficits in GS are rare and less severe phenotypes might not only be caused by mosaicism but also by less pathogenic mutations suggesting the need of a molecular genetics and functional work-up of these rare variants.
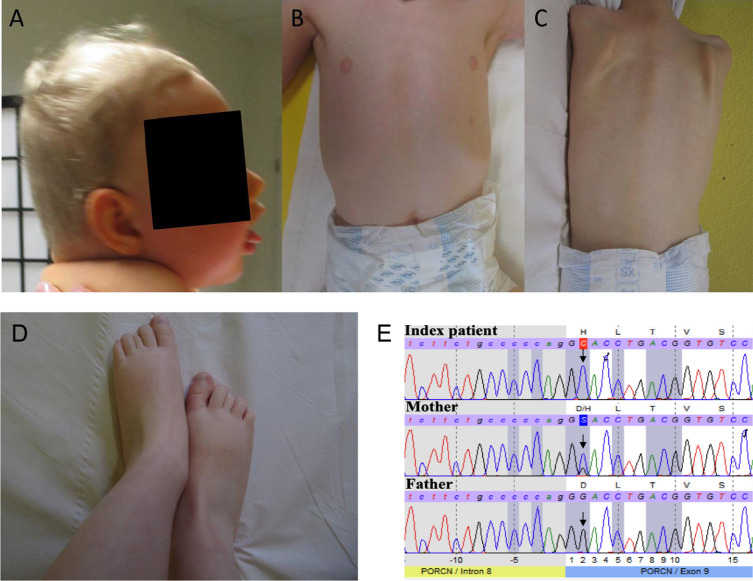
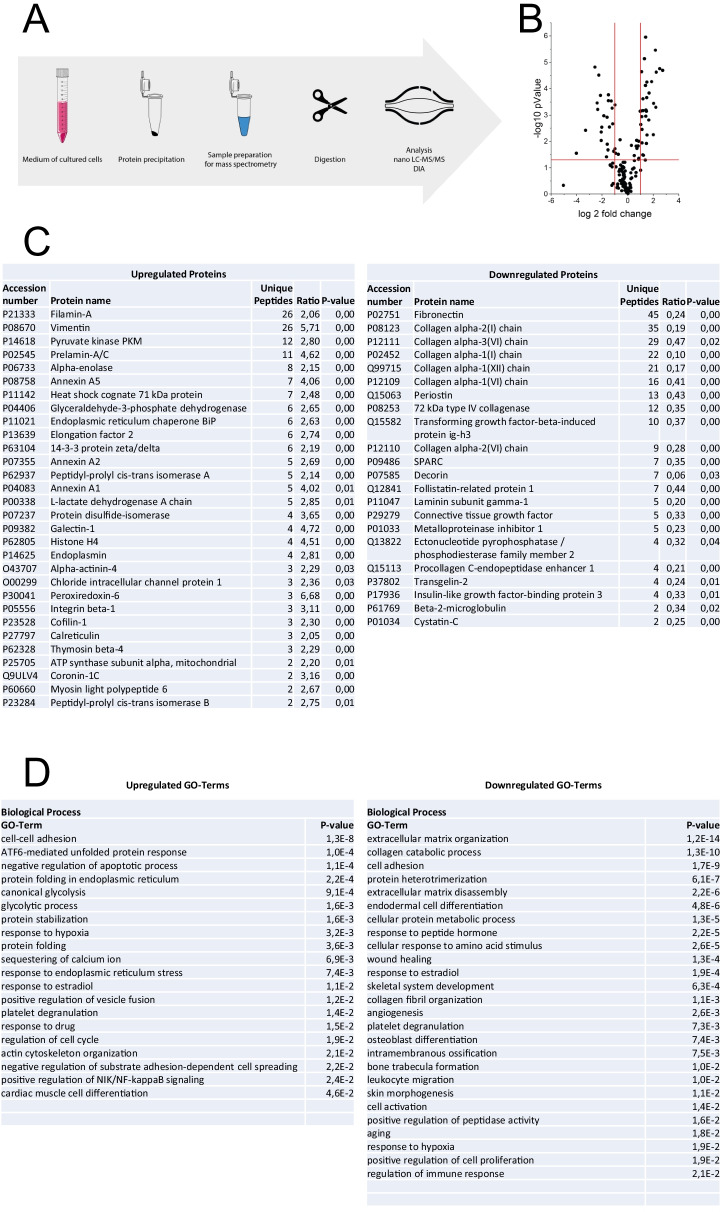
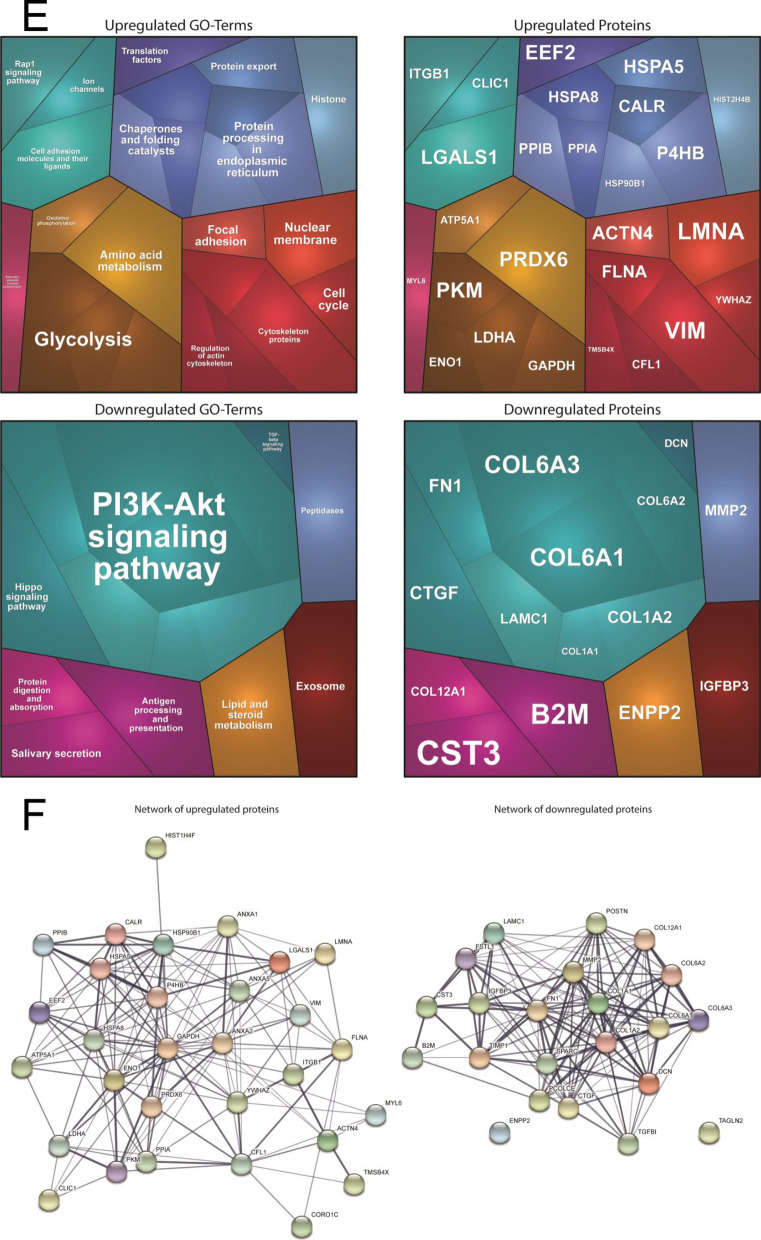
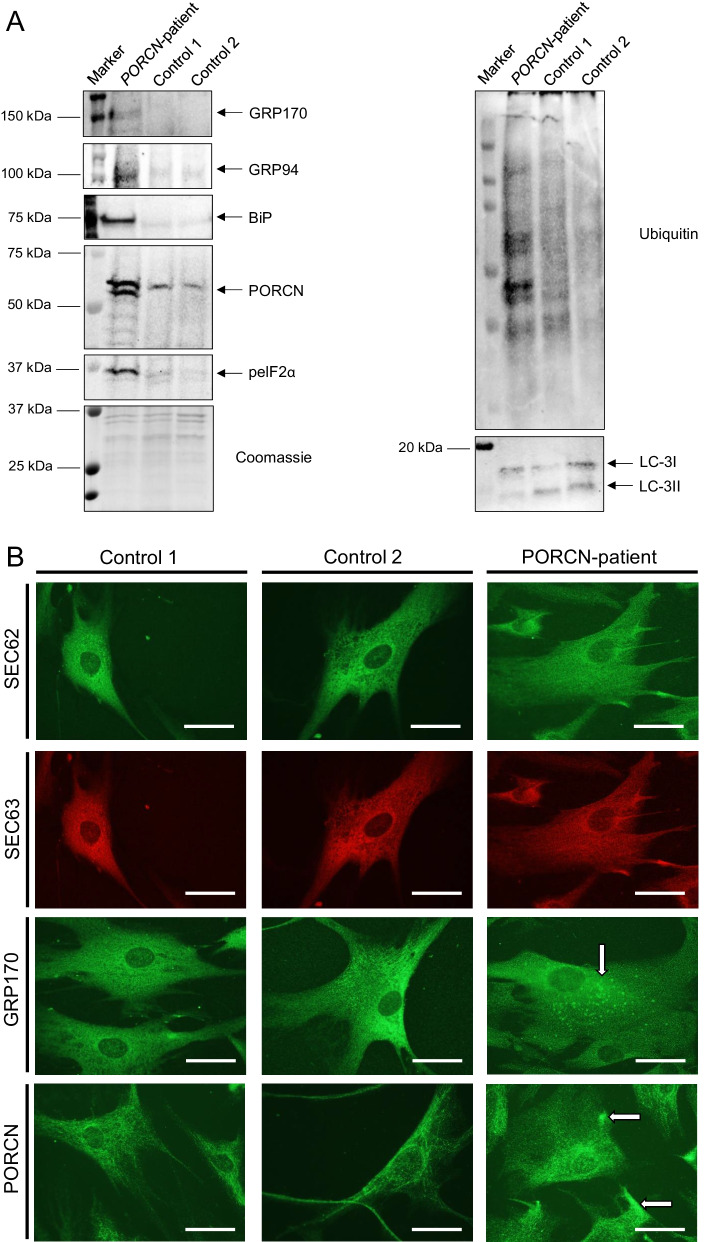
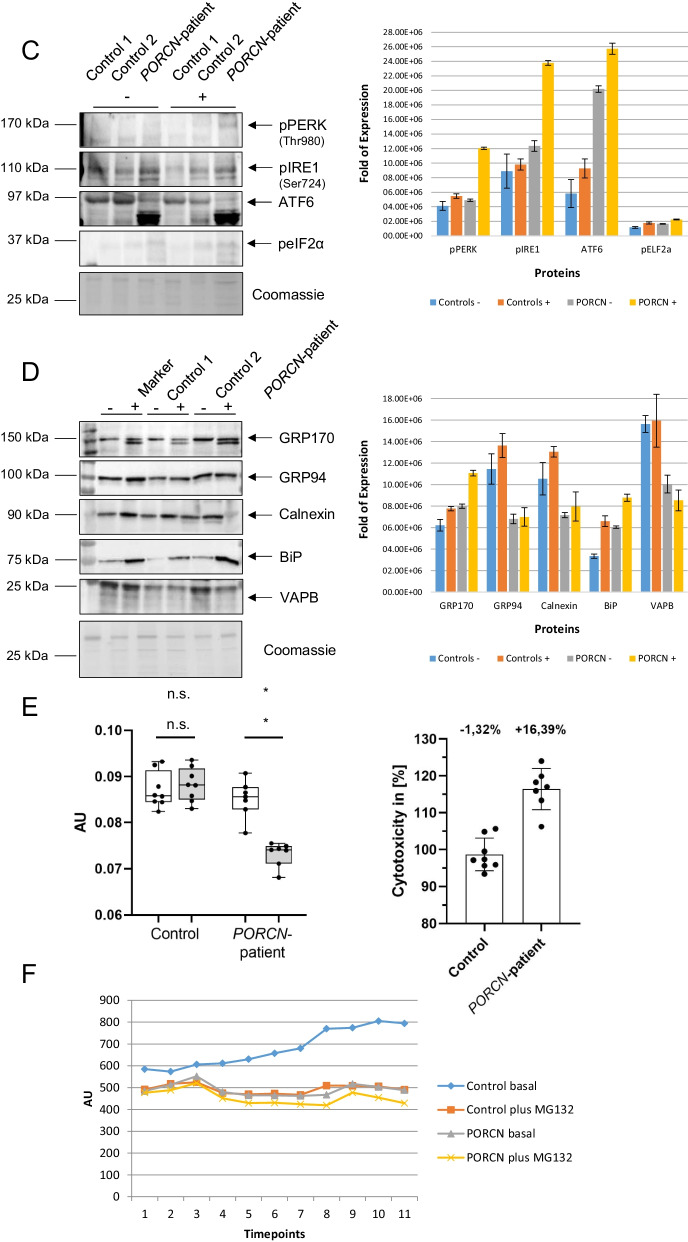
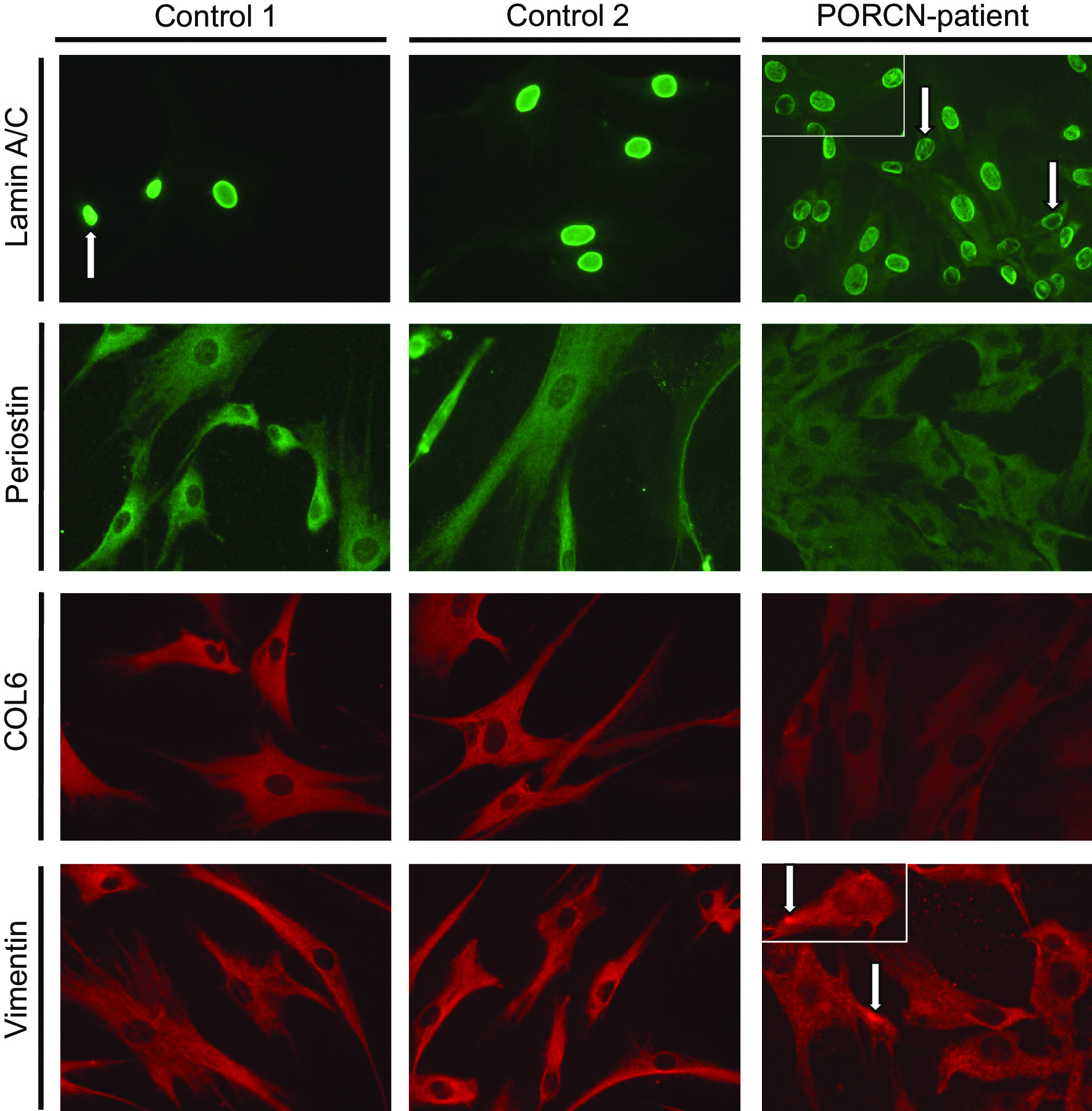
Results: We report two cases: one girl suffering from typical skin and skeletal abnormalities, developmental delay, microcephaly, thin corpus callosum, periventricular gliosis and drug-resistant epilepsy caused by a PORCN nonsense-mutation (c.283C > T, p.Arg95Ter). Presence of these combined neurological features indicates that CNS-vulnerability might be a guiding symptom in the diagnosis of GS patients. The other patient is a boy with a supernumerary nipple and skeletal anomalies but also, developmental delay, microcephaly, cerebral atrophy with delayed myelination and drug-resistant epilepsy as predominant features. Skin abnormalities were not observed. Genotyping revealed a novel PORCN missense-mutation (c.847G > C, p.Asp283His) absent in the Genome Aggregation Database (gnomAD) but also identified in his asymptomatic mother. Given that non-random X-chromosome inactivation was excluded in the mother, fibroblasts of the index had been analyzed for PORCN protein-abundance and -distribution, vulnerability against additional ER-stress burden as well as for protein secretion revealing changes.
Conclusions: Our combined findings may suggest incomplete penetrance for the p.Asp283His variant and provide novel insights into the molecular etiology of GS by adding impaired ER-function and altered protein secretion to the list of pathophysiological processes resulting in the clinical manifestation of GS.
Keywords: Connective tissue disorder; ER-stress; Fibroblast proteomics; Focal dermal hypoplasia; Goltz syndrome; Lamin a/c; Protein-serine O-palmitoleoyltransferase porcupine.
© 2022. The Author(s).
Conflict of interest statement
The authors declare not to have any competing conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
