

Atomic force microscopy reveals distinct protofilament-scale structural dynamics in depolymerizing microtubule arrays
- PMID: 35101922
- PMCID: PMC8812519
- DOI: 10.1073/pnas.2115708119
Atomic force microscopy reveals distinct protofilament-scale structural dynamics in depolymerizing microtubule arrays
Abstract
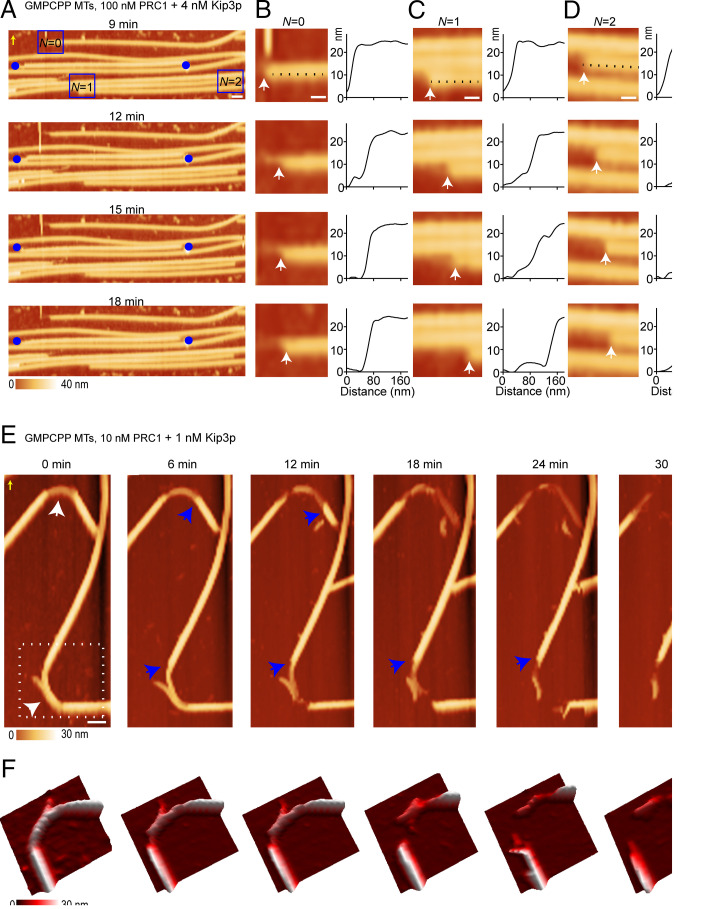
The dynamic reorganization of microtubule-based cellular structures, such as the spindle and the axoneme, fundamentally depends on the dynamics of individual polymers within multimicrotubule arrays. A major class of enzymes implicated in both the complete demolition and fine size control of microtubule-based arrays are depolymerizing kinesins. How different depolymerases differently remodel microtubule arrays is poorly understood. A major technical challenge in addressing this question is that existing optical or electron-microscopy methods lack the spatial-temporal resolution to observe the dynamics of individual microtubules within larger arrays. Here, we use atomic force microscopy (AFM) to image depolymerizing arrays at single-microtubule and protofilament resolution. We discover previously unseen modes of microtubule array destabilization by conserved depolymerases. We find that the kinesin-13 MCAK mediates asynchronous protofilament depolymerization and lattice-defect propagation, whereas the kinesin-8 Kip3p promotes synchronous protofilament depolymerization. Unexpectedly, MCAK can depolymerize the highly stable axonemal doublets, but Kip3p cannot. We propose that distinct protofilament-level activities underlie the functional dichotomy of depolymerases, resulting in either large-scale destabilization or length regulation of microtubule arrays. Our work establishes AFM as a powerful strategy to visualize microtubule dynamics within arrays and reveals how nanometer-scale substrate specificity leads to differential remodeling of micron-scale cytoskeletal structures.
Keywords: atomic force microscopy; axoneme; cytoskeleton; kinesin; microtubule arrays.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures





References
-
- Gudimchuk N. B., McIntosh J. R., Regulation of microtubule dynamics, mechanics and function through the growing tip. Nat. Rev. Mol. Cell Biol. 22, 777–795 (2021). - PubMed
-
- Kodera N., Yamamoto D., Ishikawa R., Ando T., Video imaging of walking myosin V by high-speed atomic force microscopy. Nature 468, 72–76 (2010). - PubMed
-
- Vinckier A., et al. , Immobilizing and imaging microtubules by atomic force microscopy. Ultramicroscopy 57, 337–343 (1995). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
