

The impact of storage on extracellular vesicles: A systematic study
- PMID: 35102719
- PMCID: PMC8804350
- DOI: 10.1002/jev2.12162
The impact of storage on extracellular vesicles: A systematic study
Abstract
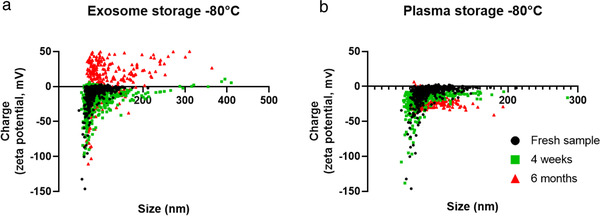
Mounting evidence suggests that storage has an impact on extracellular vesicles (EVs) properties. While -80°C storage is a widespread approach, some authors proposed improved storage strategies with conflicting results. Here, we designed a systematic study to assess the impact of -80°C storage and freeze-thaw cycles on EVs. We tested the differences among eight storage strategies and investigated the possible fusion phenomena occurring during storage. EVs were collected from human plasma and murine microglia culture by size exclusion chromatography and ultracentrifugation, respectively. The analysis included: concentration, size and zeta potential (tunable resistive pulse sensing), contaminant protein assessment; flow cytometry for the analysis of two single fluorescent-tagged EVs populations (GFP and mCherry), mixed before preservation. We found that -80°C storage reduces EVs concentration and sample purity in a time-dependent manner. Furthermore, it increases the particle size and size variability and modifies EVs zeta potential, with a shift of EVs in size-charge plots. None of the tested conditions prevented the observed effects. Freeze-thaw cycles lead to an EVs reduction after the first cycle and to a cycle-dependent increase in particle size. With flow cytometry, after storage, we observed a significant population of double-positive EVs (GFP+ -mCherry+ ). This observation may suggest the occurrence of fusion phenomena during storage. Our findings show a significant impact of storage on EVs samples in terms of particle loss, purity reduction and fusion phenomena leading to artefactual particles. Depending on downstream analyses and experimental settings, EVs should probably be processed from fresh, non-archival, samples in majority of cases.
Keywords: extracellular vesicles; flow cytometry; fusion; preservation; size exclusion chromatography; storage; tunable resistive pulse sensing.
© 2022 The Authors. Journal of Extracellular Vesicles published by Wiley Periodicals, LLC on behalf of the International Society for Extracellular Vesicles.
Conflict of interest statement
Gelibter S, Marostica G, Mandelli A, Siciliani S, Podini P, Finardi A report no disclosures. Furlan R received honoraria for serving on scientifc advisory boards or as a speaker from Biogen, Novartis, Roche, Merck, and funding for research from Merck.
Figures






References
-
- Agrawal, A. K. , Aqil, F. , Jeyabalan, J. , Spencer, W. A. , Beck, J. , Gachuki, B. W. , Alhakeem, S. S. , Oben, K. , Munagala, R. , Bondada, S. , & Gupta, R. C. (2017).Milk‐derived exosomes for oral delivery of paclitaxel. Nanomedicine: Nanotechnology, Biology and Medicine, 13(5), 1627–1636. - PubMed
-
- Ayers, L. , Kohler, M. , Harrison, P. , Sargent, I. , Dragovic, R. , Schaap, M. , Nieuwland, R. , Brooks, S. A. , & Ferry, B. (2011).Measurement of circulating cell‐derived microparticles by flow cytometry: Sources of variability within the assay. Thrombosis Research, 127(4), 370–377. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
