

Host Cellular RNA Helicases Regulate SARS-CoV-2 Infection
- PMID: 35107372
- PMCID: PMC8941876
- DOI: 10.1128/jvi.00002-22
Host Cellular RNA Helicases Regulate SARS-CoV-2 Infection
Abstract
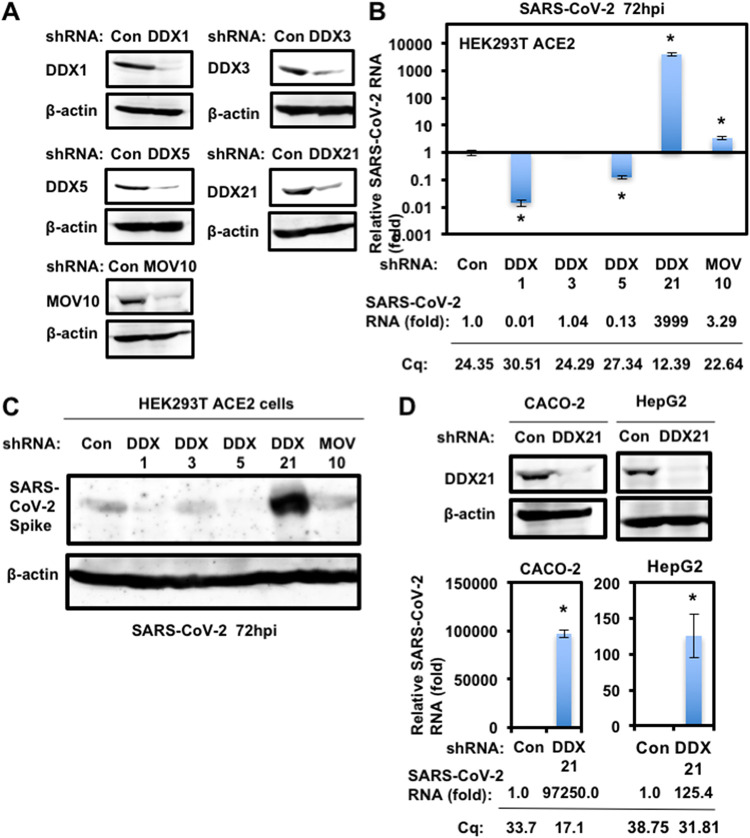
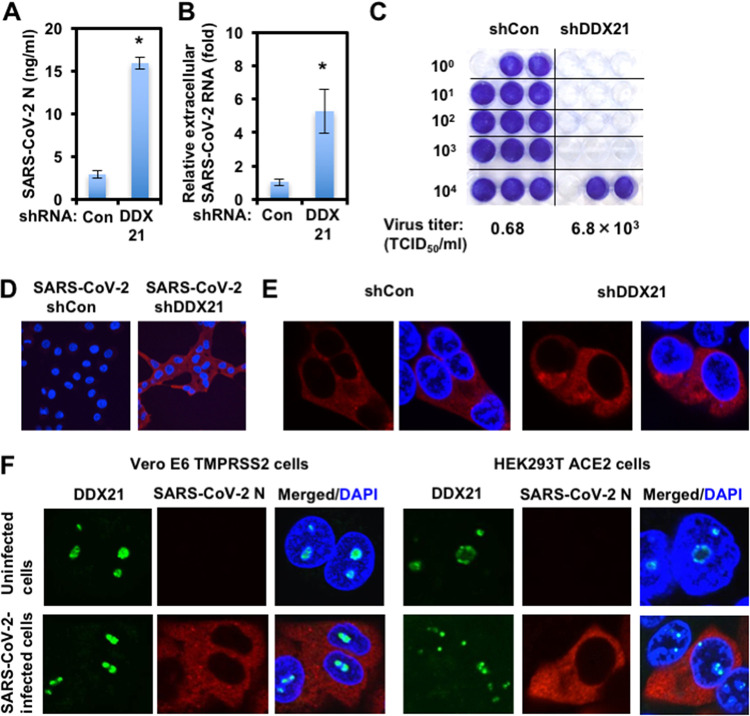
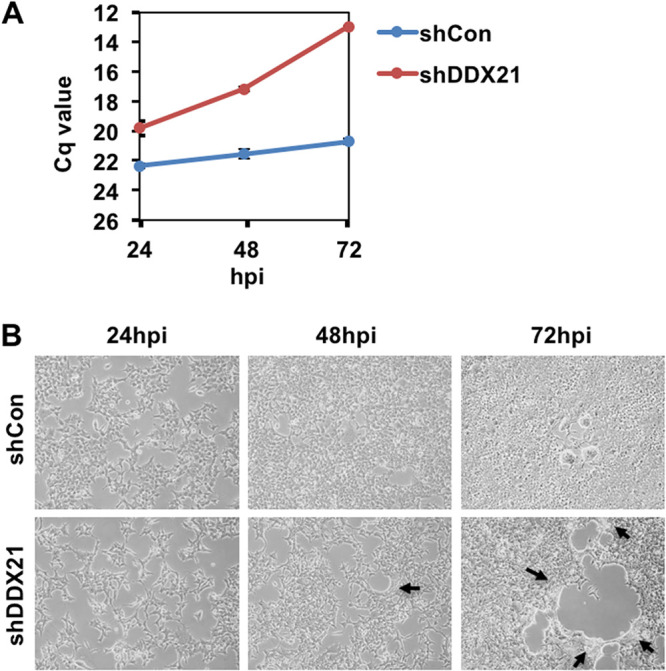
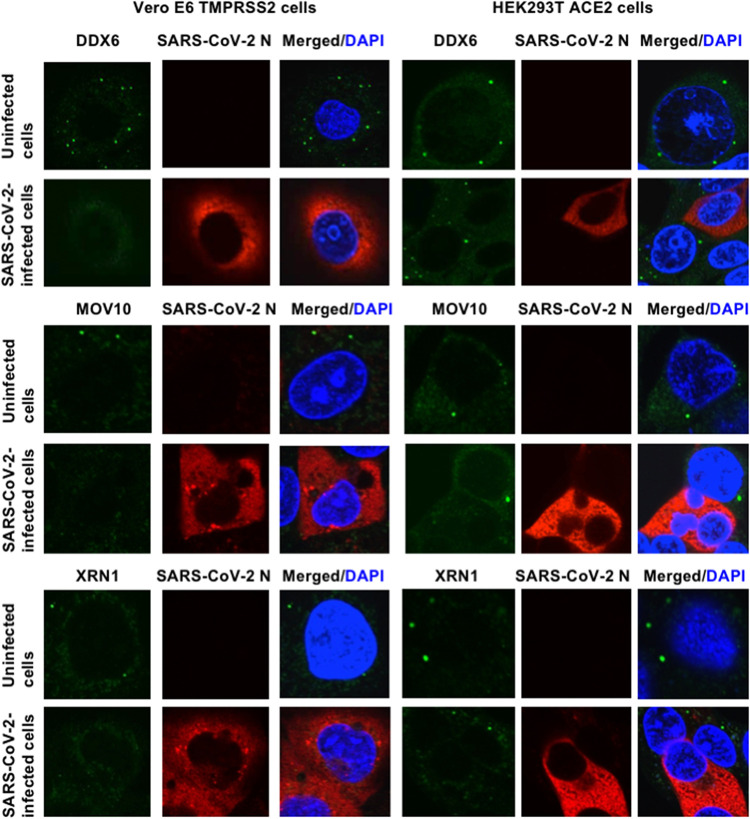
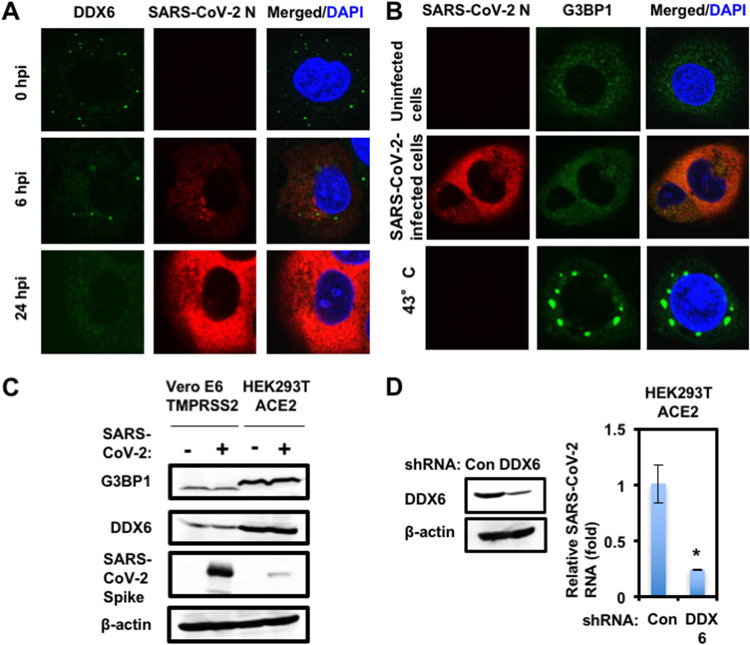
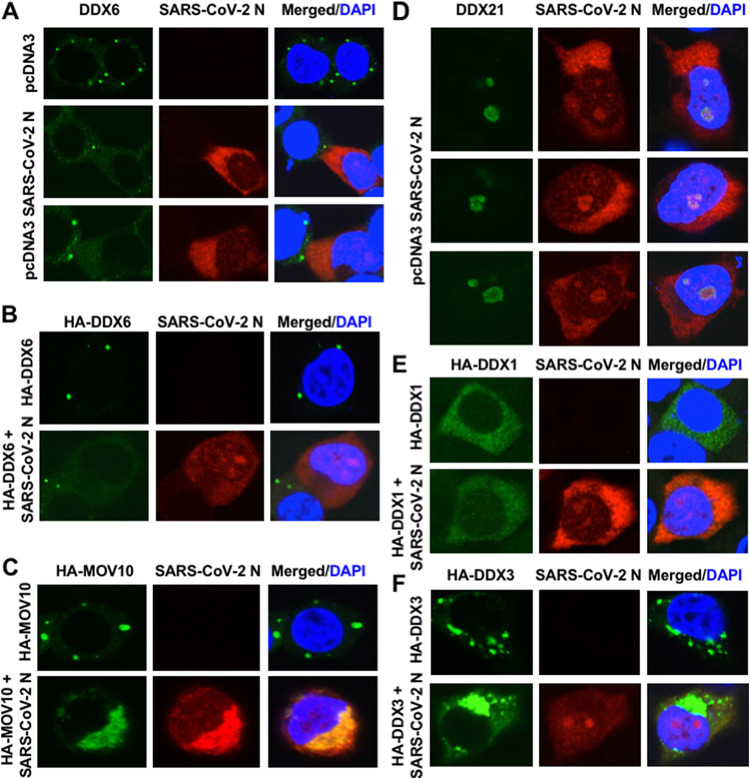
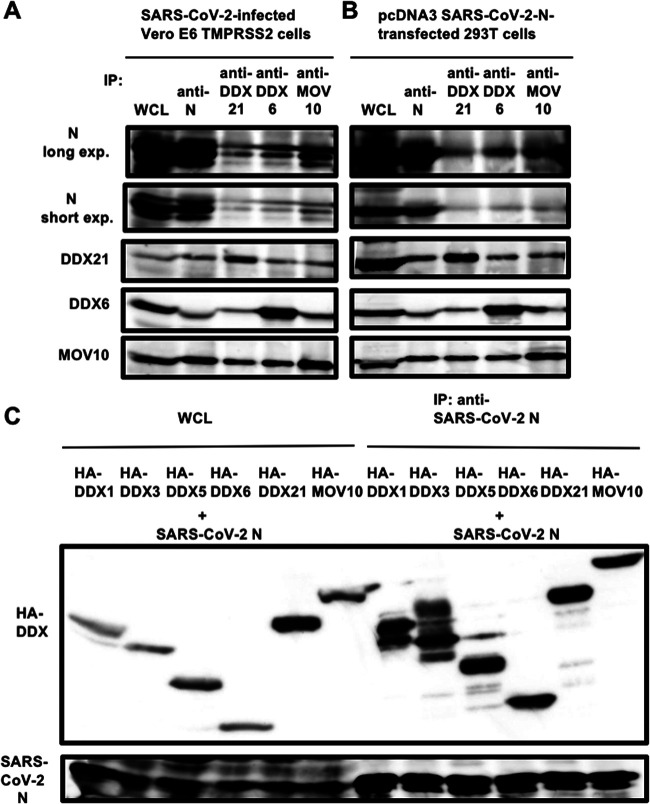
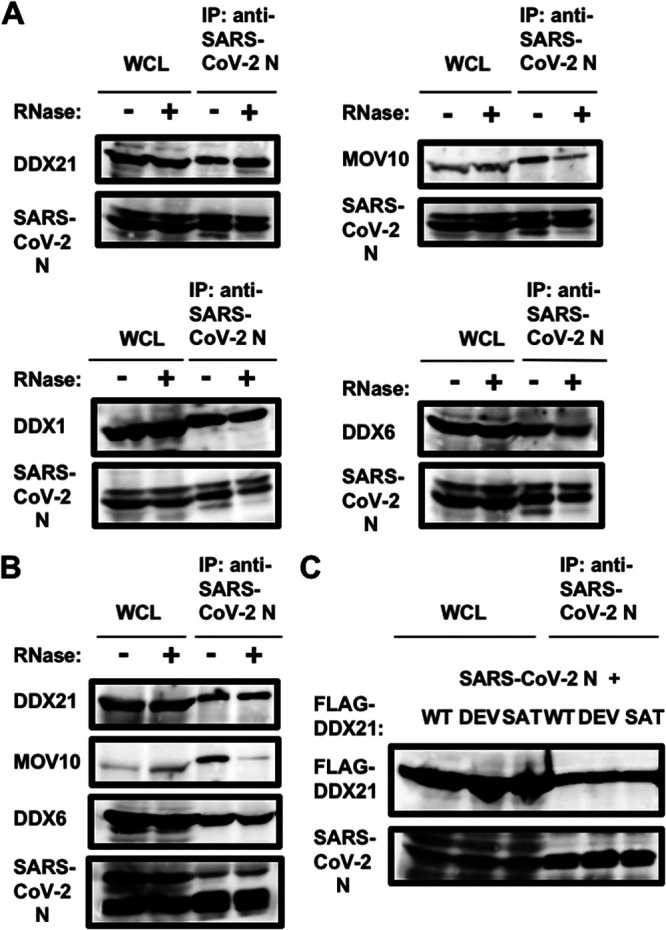
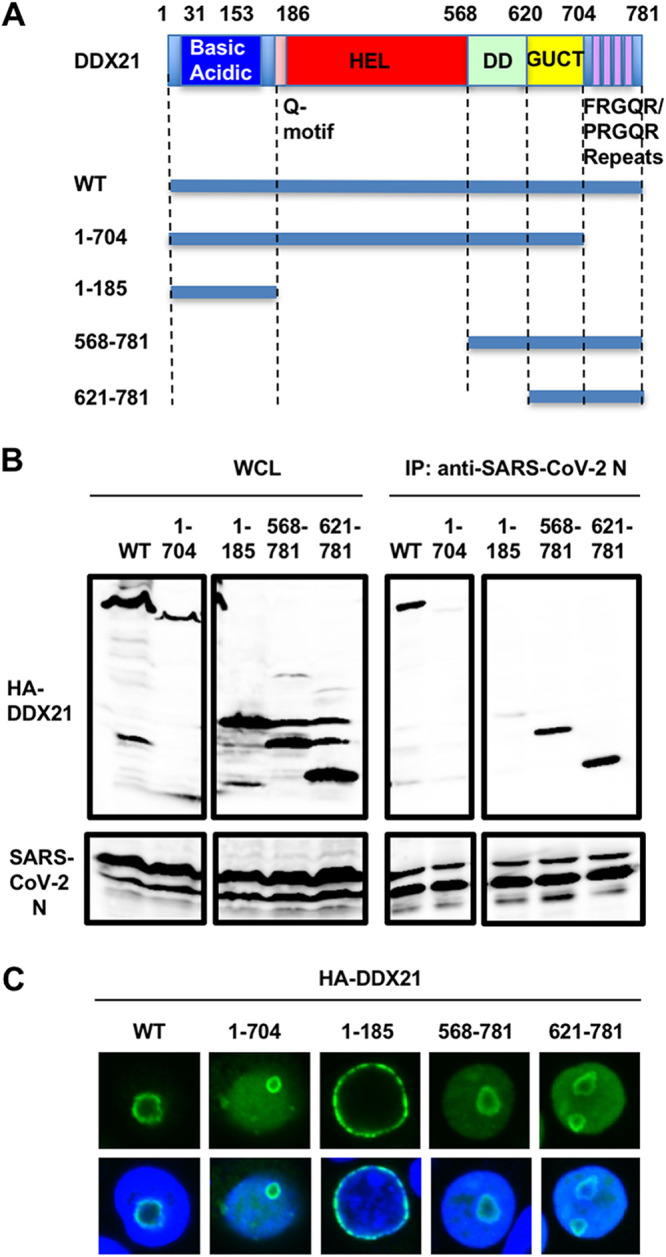
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) has the largest RNA genome, approximately 30 kb, among RNA viruses. The DDX DEAD box RNA helicase is a multifunctional protein involved in all aspects of RNA metabolism. Therefore, host RNA helicases may regulate and maintain such a large viral RNA genome. In this study, I investigated the potential role of several host cellular RNA helicases in SARS-CoV-2 infection. Notably, DDX21 knockdown markedly accumulated intracellular viral RNA and viral production, as well as viral infectivity of SARS-CoV-2, indicating that DDX21 strongly restricts the SARS-CoV-2 infection. In addition, MOV10 RNA helicase also suppressed the SARS-CoV-2 infection. In contrast, DDX1, DDX5, and DDX6 RNA helicases were required for SARS-CoV-2 replication. Indeed, SARS-CoV-2 infection dispersed the P-body formation of DDX6 and MOV10 RNA helicases as well as XRN1 exonuclease, while the viral infection did not induce stress granule formation. Accordingly, the SARS-CoV-2 nucleocapsid (N) protein interacted with DDX1, DDX3, DDX5, DDX6, DDX21, and MOV10 and disrupted the P-body formation, suggesting that SARS-CoV-2 N hijacks DDX6 to carry out viral replication. Conversely, DDX21 and MOV10 restricted SARS-CoV-2 infection through an interaction of SARS-CoV-2 N with host cellular RNA helicases. Altogether, host cellular RNA helicases seem to regulate the SARS-CoV-2 infection. IMPORTANCE SARS-CoV-2 has a large RNA genome, of approximately 30 kb. To regulate and maintain such a large viral RNA genome, host RNA helicases may be involved in SARS-CoV-2 replication. In this study, I have demonstrated that DDX21 and MOV10 RNA helicases limit viral infection and replication. In contrast, DDX1, DDX5, and DDX6 are required for SARS-CoV-2 infection. Interestingly, SARS-CoV-2 infection disrupted P-body formation and attenuated or suppressed stress granule formation. Thus, SARS-CoV-2 seems to hijack host cellular RNA helicases to play a proviral role by facilitating viral infection and replication and by suppressing the host innate immune system.
Keywords: DDX1; DDX21; DDX6; P-body; RNA helicase; SARS-CoV-2; coronavirus; nucleocapsid; nucleolus; stress granule.
Conflict of interest statement
The authors declare no conflict of interest.
We have no conflicts of interest to disclose.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
