

Neuroendocrine control of gonadotropin-releasing hormone: Pulsatile and surge modes of secretion
- PMID: 35107859
- PMCID: PMC9948945
- DOI: 10.1111/jne.13094
Neuroendocrine control of gonadotropin-releasing hormone: Pulsatile and surge modes of secretion
Abstract
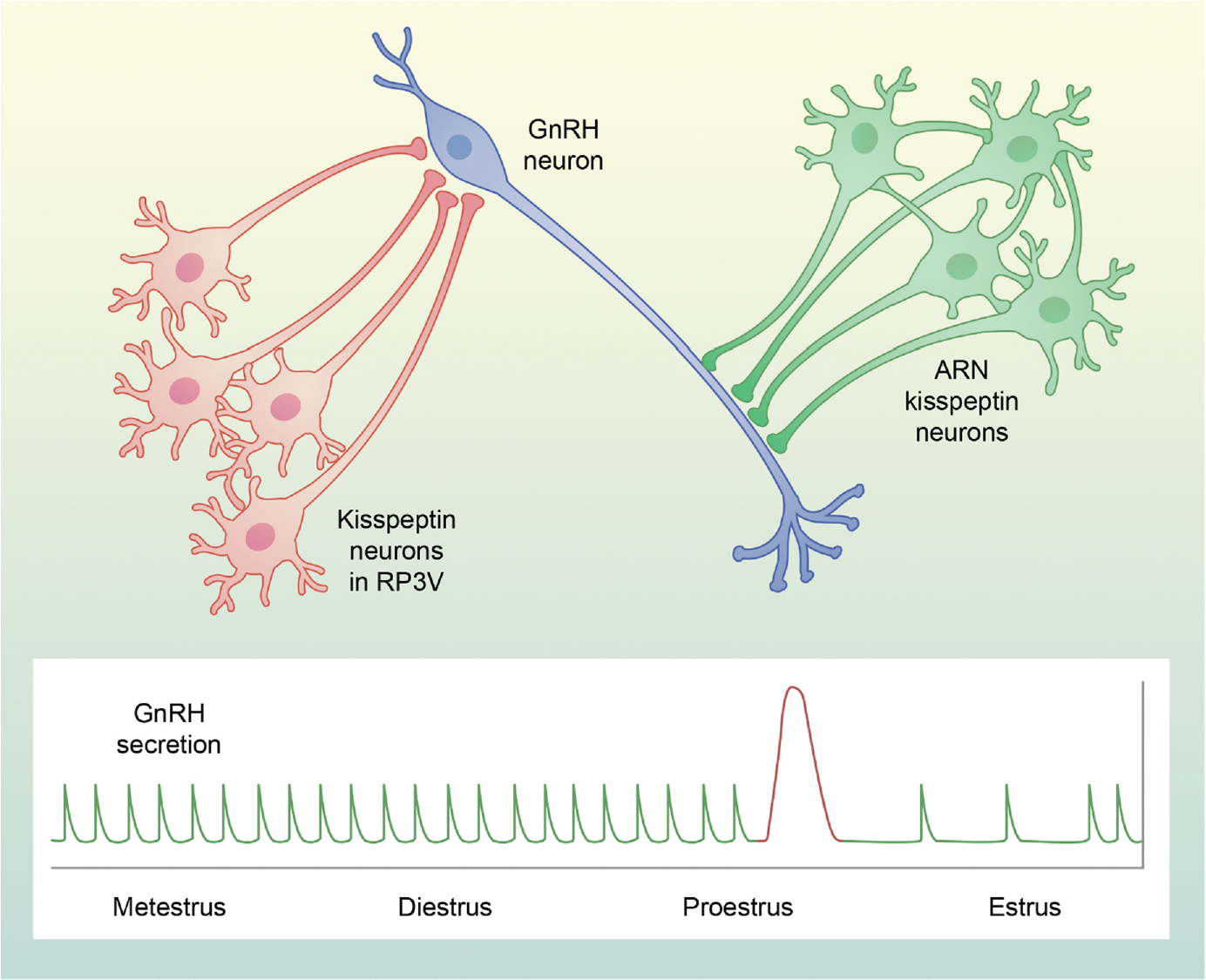
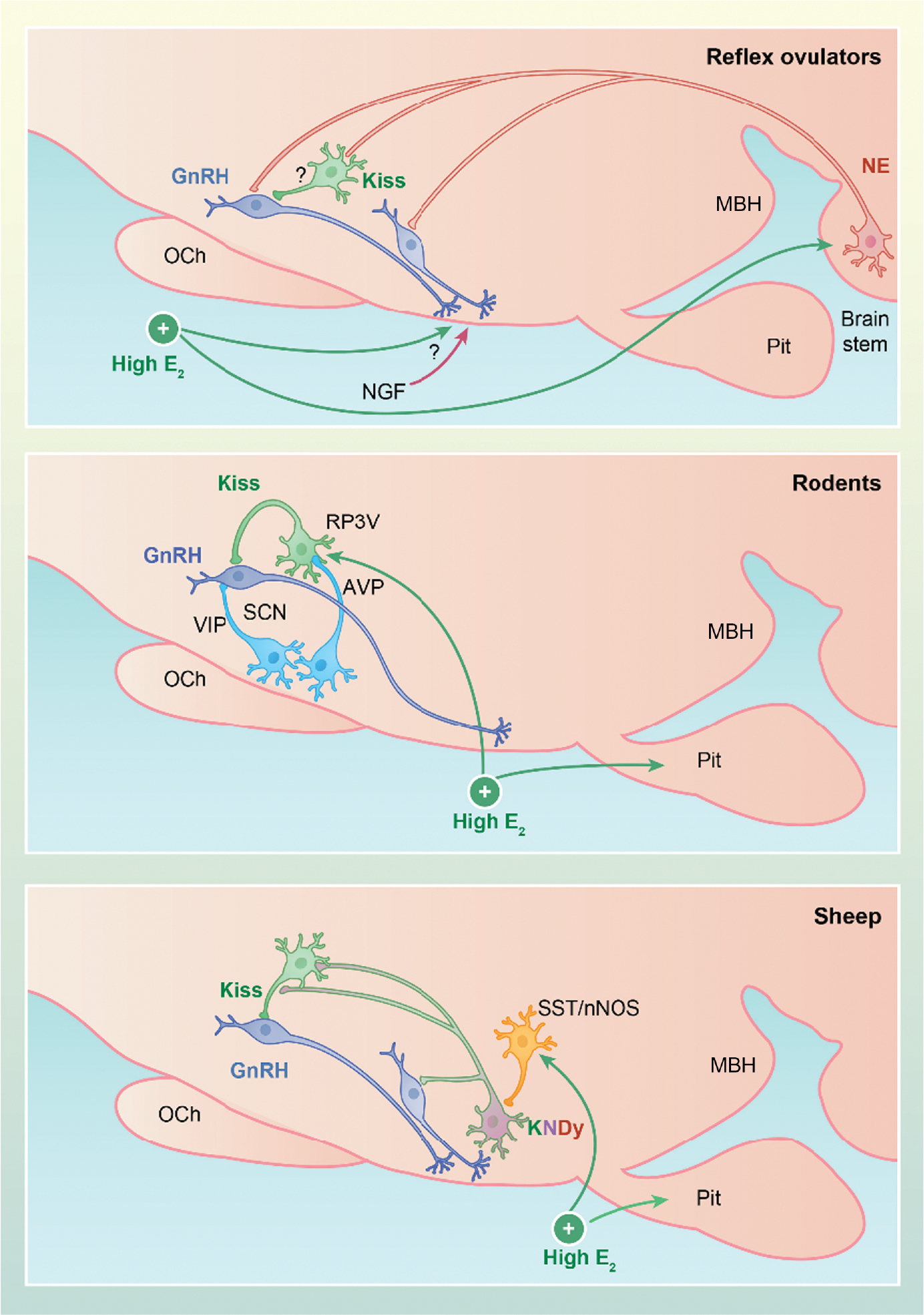
The concept that different systems control episodic and surge secretion of gonadotropin-releasing hormone (GnRH) was well established by the time that GnRH was identified and formed the framework for studies of the physiological roles of GnRH, and later kisspeptin. Here, we focus on recent studies identifying the neural mechanisms underlying these two modes of secretion, with an emphasis on their core components. There is now compelling data that kisspeptin neurons in the arcuate nucleus that also contain neurokinin B (NKB) and dynorphin (i.e., KNDy cells) and their projections to GnRH dendrons constitute the GnRH pulse generator in mice and rats. There is also strong evidence for a similar role for KNDy neurons in sheep and goats, and weaker data in monkeys and humans. However, whether KNDy neurons act on GnRH dendrons and/or GnRH soma and dendrites that are found in the mediobasal hypothalamus (MBH) of these species remains unclear. The core components of the GnRH/luteinising hormone surge consist of an endocrine signal that initiates the process and a neural trigger that drives GnRH secretion during the surge. In all spontaneous ovulators, the core endocrine signal is a rise in estradiol secretion from the maturing follicle(s), with the site of estrogen positive feedback being the rostral periventricular kisspeptin neurons in rodents and neurons in the MBH of sheep and primates. There is considerable species variations in the neural trigger, with three major classes. First, in reflex ovulators, this trigger is initiated by coitus and carried to the hypothalamus by neural or vascular pathways. Second, in rodents, there is a time of day signal that originates in the suprachiasmatic nucleus and activates rostral periventricular kisspeptin neurons and GnRH soma and dendrites. Finally, in sheep nitric oxide-producing neurons in the ventromedial nucleus, KNDy neurons and rostral kisspeptin neurons all appear to participate in driving GnRH release during the surge.
Keywords: GnRH pulses; GnRH surge; KNDy neurons; kisspeptin; steroid feedback.
© 2022 British Society for Neuroendocrinology.
Conflict of interest statement
CONFLICT OF INTERESTS
The authors declare that they have no conflicts of interest.
Figures
References
-
- Everett JW, Sawyer CH, Markee JE. A neurogenic timing factor in control of the ovulatory discharge of luteinizing hormone in the cyclic rat. Endocrinology. 1949;44(3):234–250. - PubMed
-
- Halasz B, Gorski RA. Gonadotrophic hormone secretion in female rats after partial or total interruption of neural afferents to the medial basal hypothalamus. Endocrinology. 1967;80(4):608–622. - PubMed
-
- Dungan HM, Clifton DK, Steiner RA. Minireview: kisspeptin neurons as central processors in the regulation of gonadotropin-releasing hormone secretion. Endocrinology. 2006;147(3):1154–1158. - PubMed
-
- Dierschke DJ, Bhattacharya AN, Atkinson LE, Knobil E. Circhoral oscillations of plasma LH levels in the ovariectomized rhesus monkey. Endocrinology. 1970;87(5):850–853. - PubMed
-
- Schally AV, Arimura A, Baba Y, et al. Isolation and properties of the FSH and LH-releasing hormone. Biochem Biophys Res Comm. 1971;43(2):393–399. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
