

Structure-guided stabilization of pathogen-derived peptide-HLA-E complexes using non-natural amino acids conserves native TCR recognition
- PMID: 35108401
- PMCID: PMC9306587
- DOI: 10.1002/eji.202149745
Structure-guided stabilization of pathogen-derived peptide-HLA-E complexes using non-natural amino acids conserves native TCR recognition
Abstract
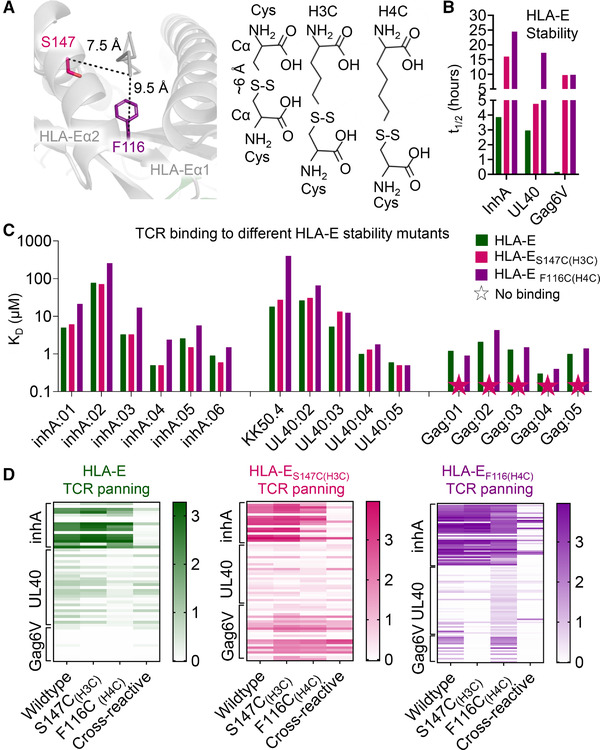
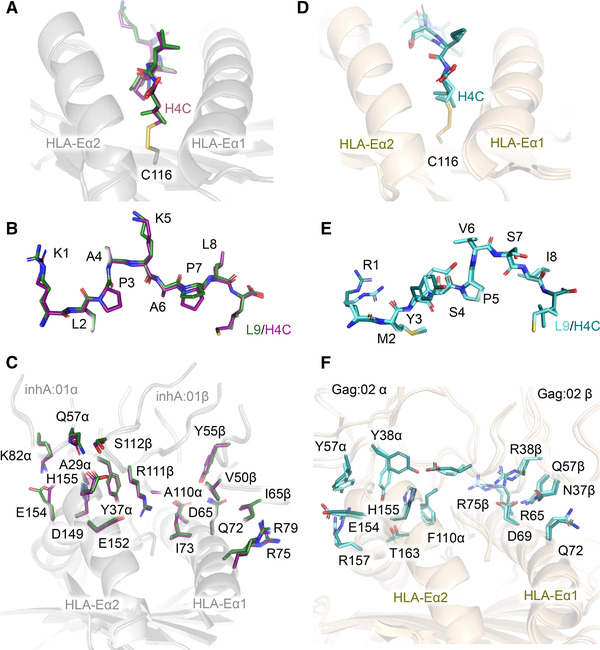
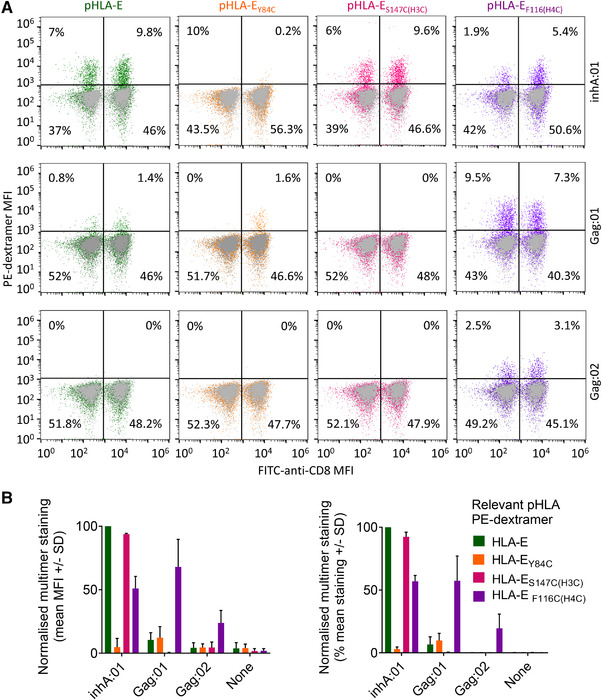
The nonpolymorphic class Ib molecule, HLA-E, primarily presents peptides from HLA class Ia leader peptides, providing an inhibitory signal to NK cells via CD94/NKG2 interactions. Although peptides of pathogenic origin can also be presented by HLA-E to T cells, the molecular basis underpinning their role in antigen surveillance is largely unknown. Here, we solved a co-complex crystal structure of a TCR with an HLA-E presented peptide (pHLA-E) from bacterial (Mycobacterium tuberculosis) origin, and the first TCR-pHLA-E complex with a noncanonically presented peptide from viral (HIV) origin. The structures provided a molecular foundation to develop a novel method to introduce cysteine traps using non-natural amino acid chemistry that stabilized pHLA-E complexes while maintaining native interface contacts between the TCRs and different pHLA-E complexes. These pHLA-E monomers could be used to isolate pHLA-E-specific T cells, with obvious utility for studying pHLA-E restricted T cells, and for the identification of putative therapeutic TCRs.
Keywords: HIV; HLA-E; TCR; crystal structure; non-natural amino acids.
© 2022 Immunocore. European Journal of Immunology published by Wiley-VCH GmbH.
Conflict of interest statement
All authors are, or were, employees of Immunocore Ltd.
Figures
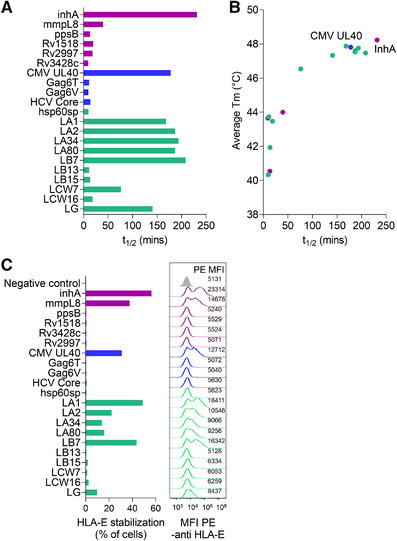
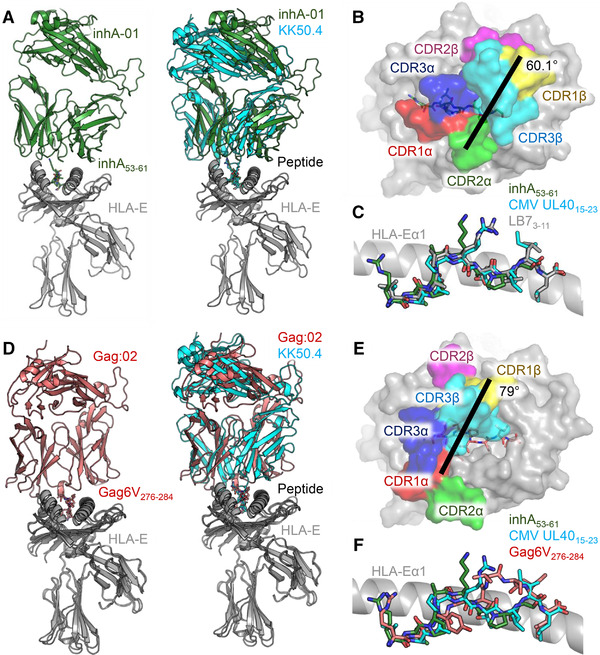
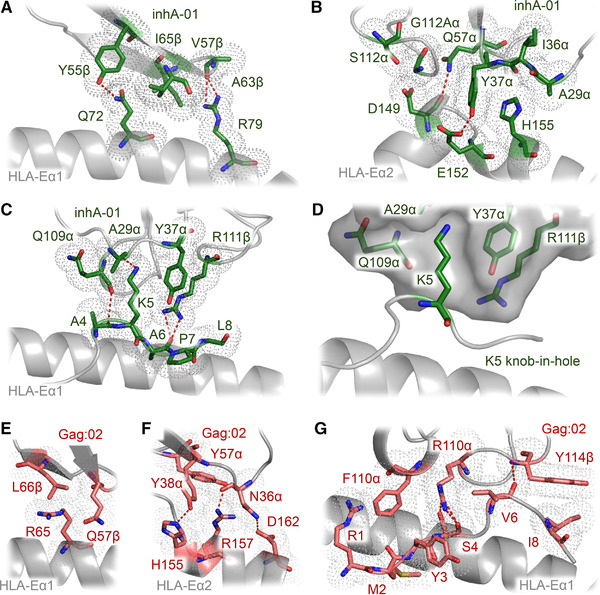
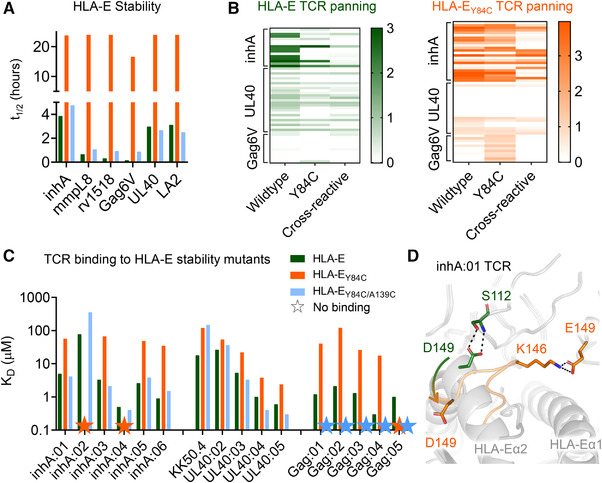
References
-
- Geraghty, D. E. , Stockschleader, M. , Ishitani, A. and Hansen, J. A. , Polymorphism at the HLA‐E locus predates most HLA‐A and ‐B polymorphism. Hum. Immunol. 1992. 33: 174–184. - PubMed
-
- Tomasec, P. , Braud, V. M. , Rickards, C. , Powell, M. B. , McSharry, B. P. , Gadola, S. , Cerundolo, V. et al., Surface expression of HLA‐E, an inhibitor of natural killer cells, enhanced by human cytomegalovirus gpUL40. Science (New York, N.Y.). 2000. 287: 1031. - PubMed
-
- Grimsley, C. and Ober, C. , Population genetic studies of HLA‐E. Hum. Immunol. 1997. 52: 33–40. - PubMed
-
- Strong, R. K. , Holmes, M. A. , Li, P. , Braun, L. , Lee, N. and Geraghty, D. E. , HLA‐E allelic variants. J. Biol. Chem. 2003. 278: 5082–5090. - PubMed
-
- Braud, V. , Yvonne Jones, E. and McMichael, A. , The human major histocompatibility complex class Ib molecule HLA‐E binds signal sequence‐derived peptides with primary anchor residues at positions 2 and 9. Eur. J. Immunol. . 1997. 27: 1164–1169. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
