

Heterogenous impairment of α cell function in type 2 diabetes is linked to cell maturation state
- PMID: 35108513
- PMCID: PMC8852281
- DOI: 10.1016/j.cmet.2021.12.021
Heterogenous impairment of α cell function in type 2 diabetes is linked to cell maturation state
Abstract
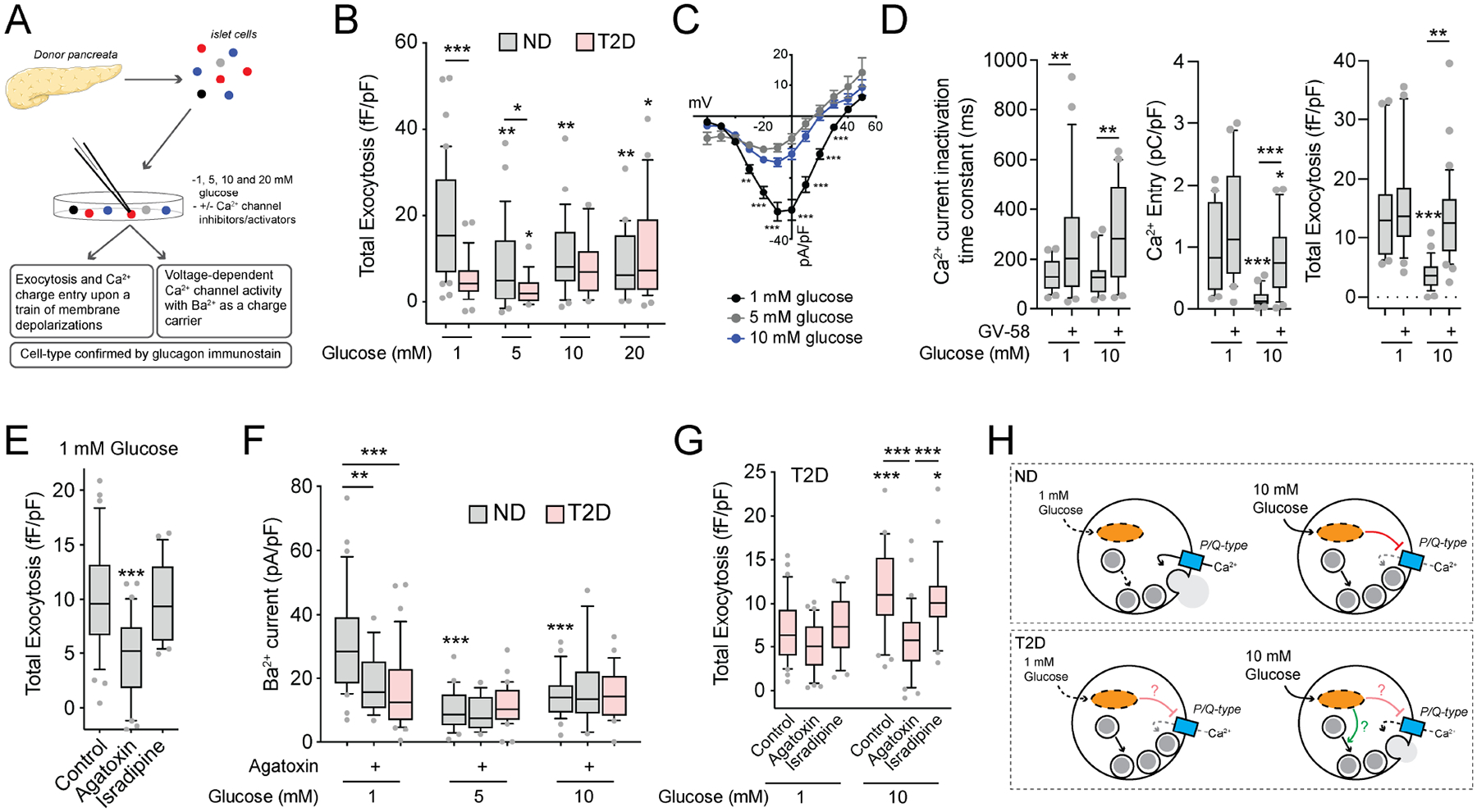
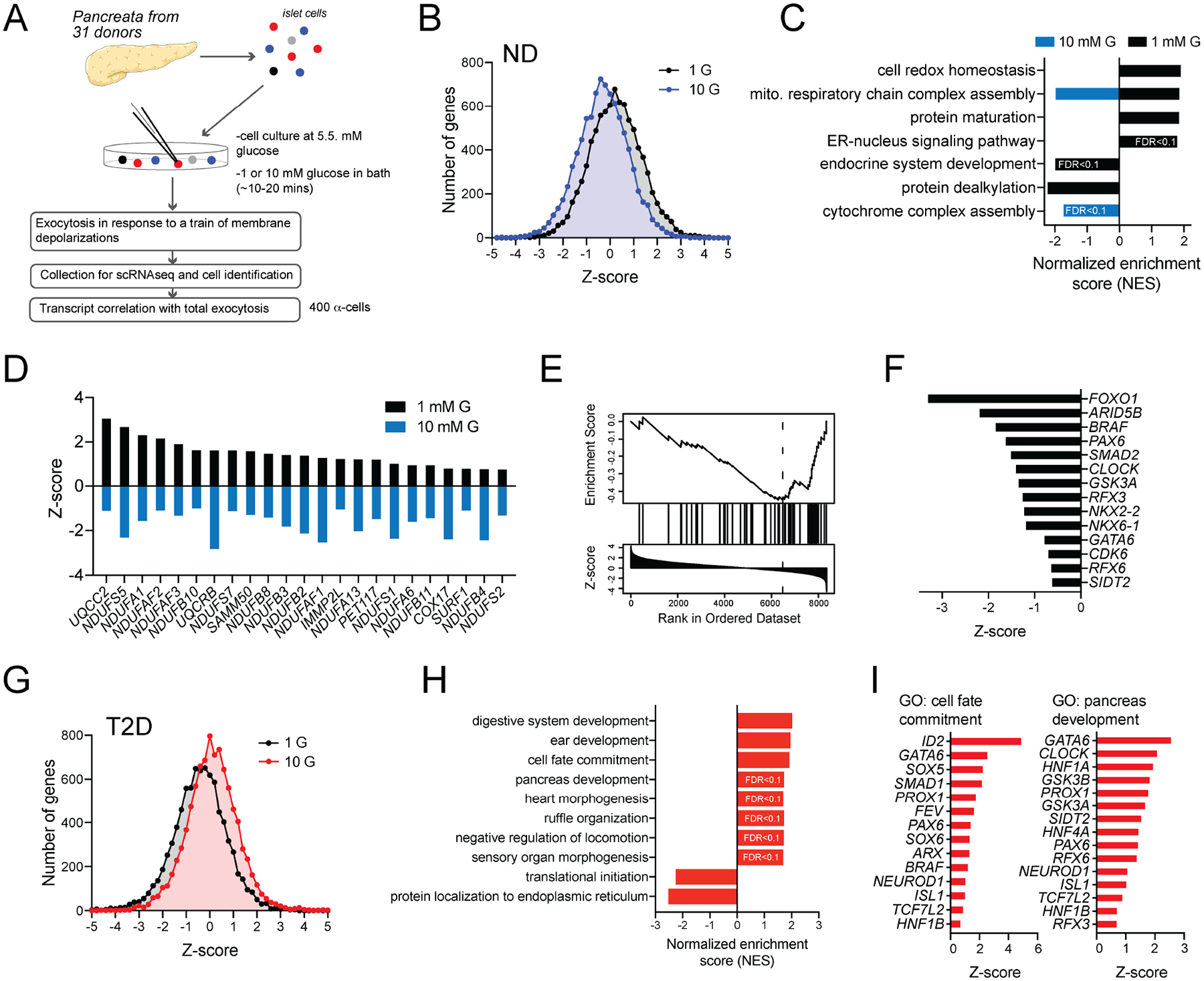
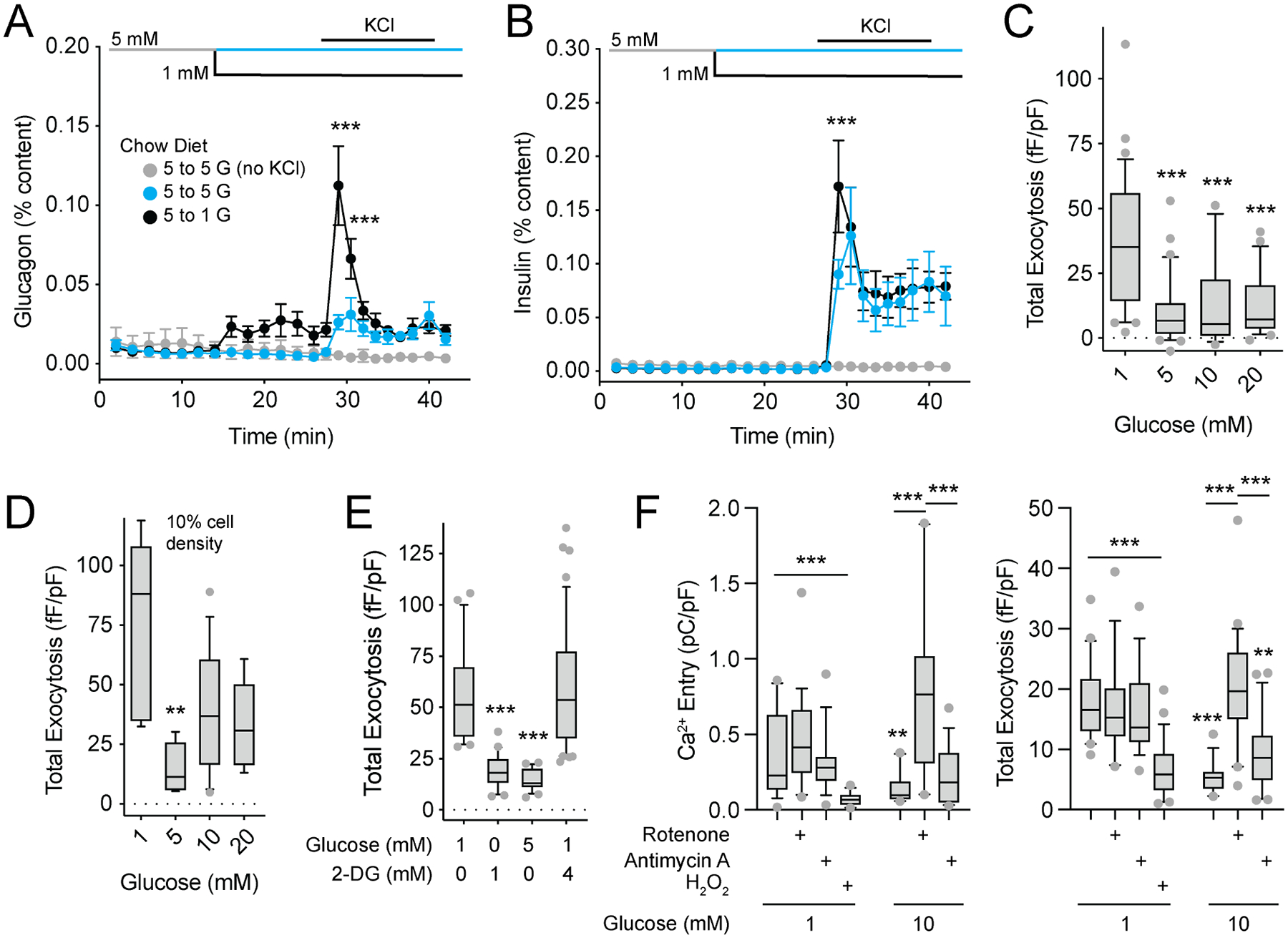
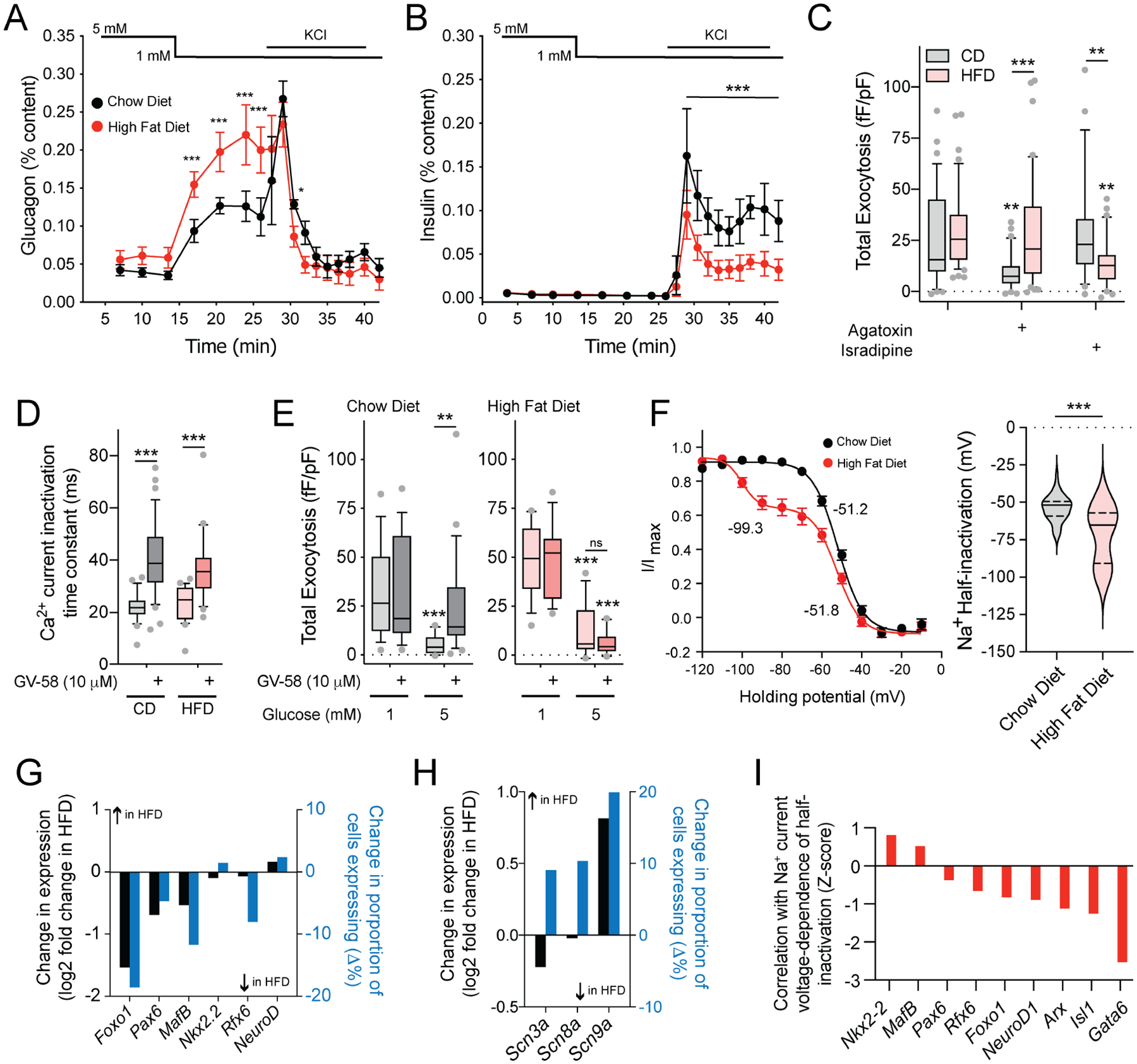
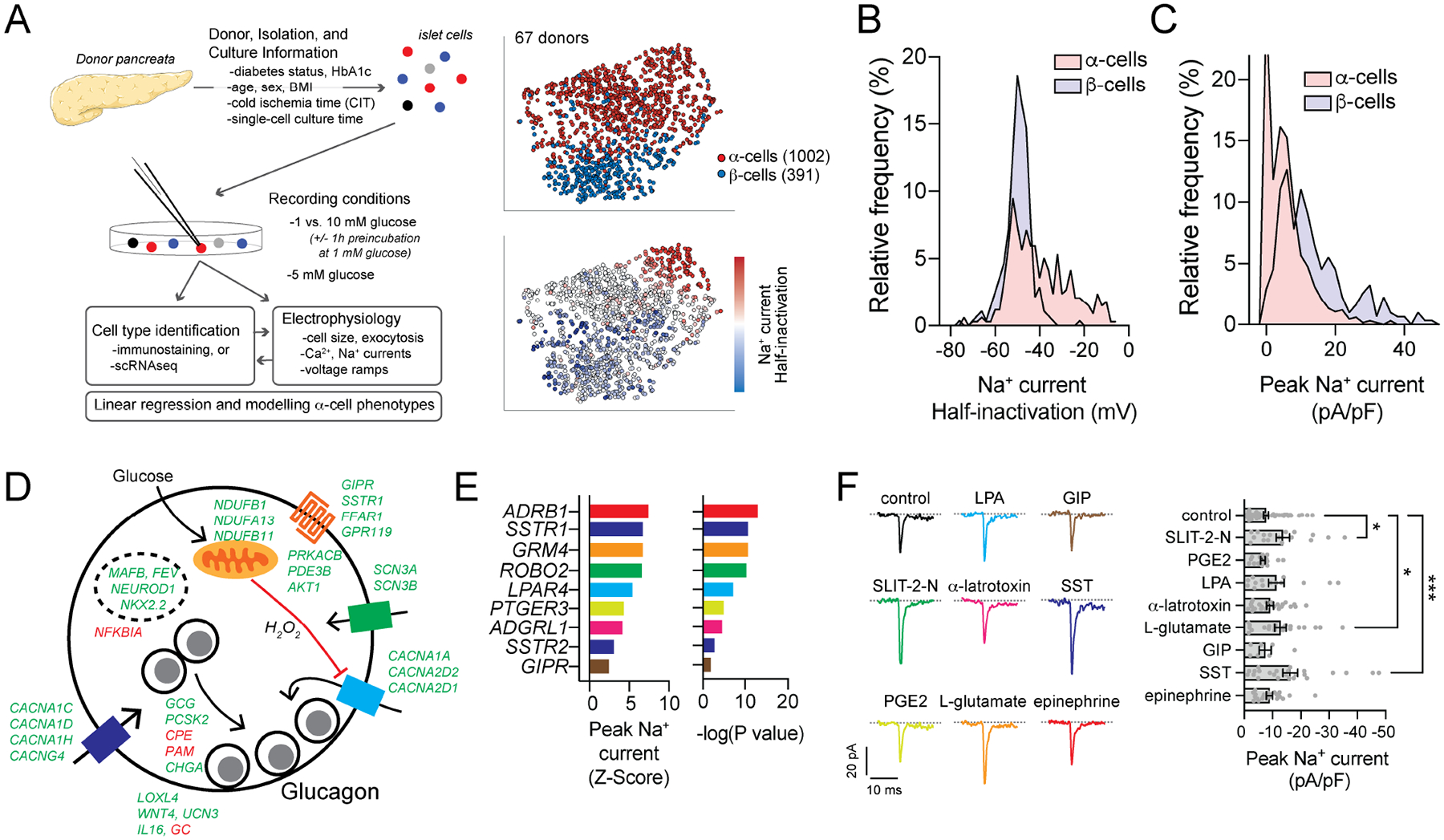
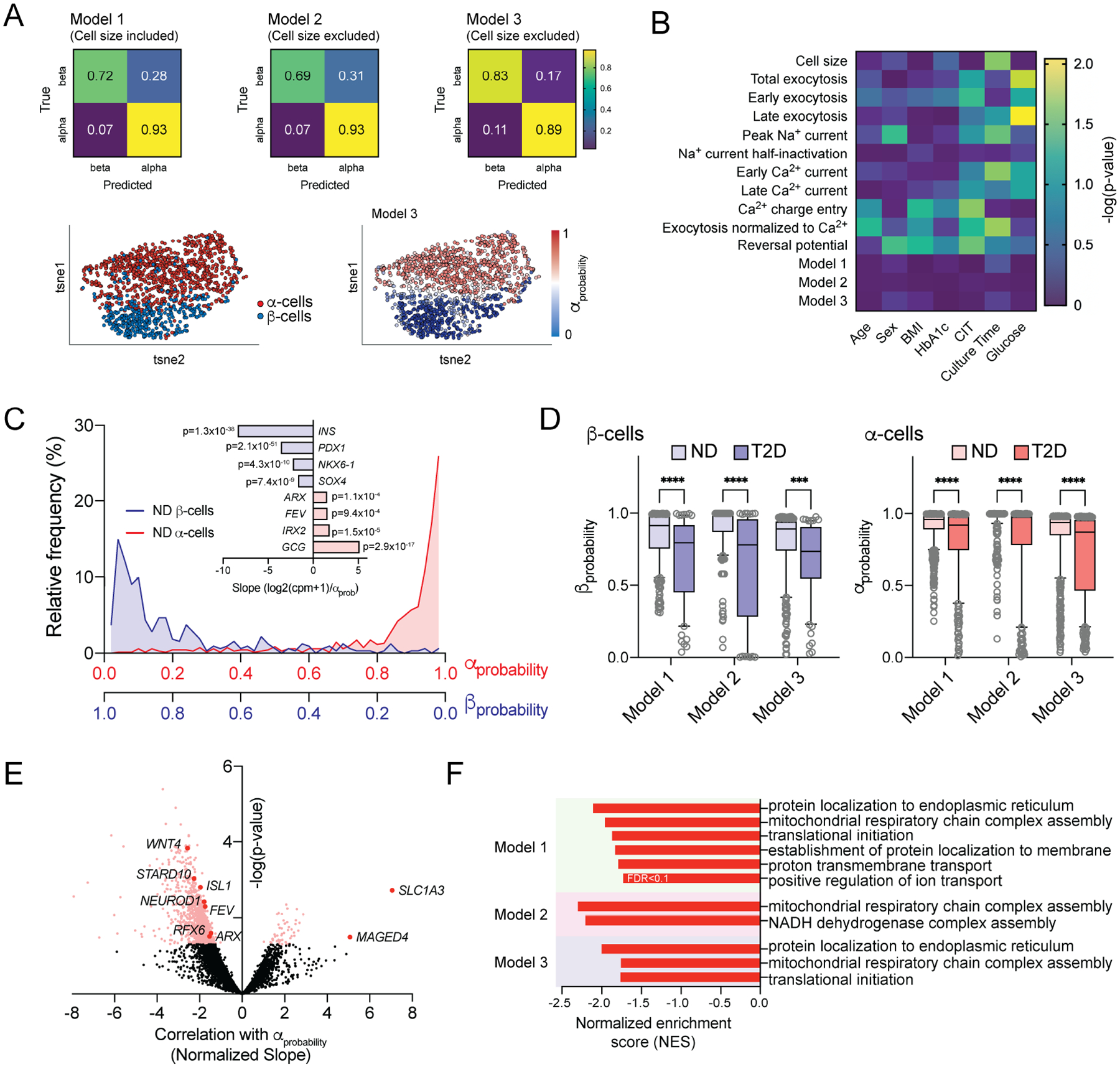
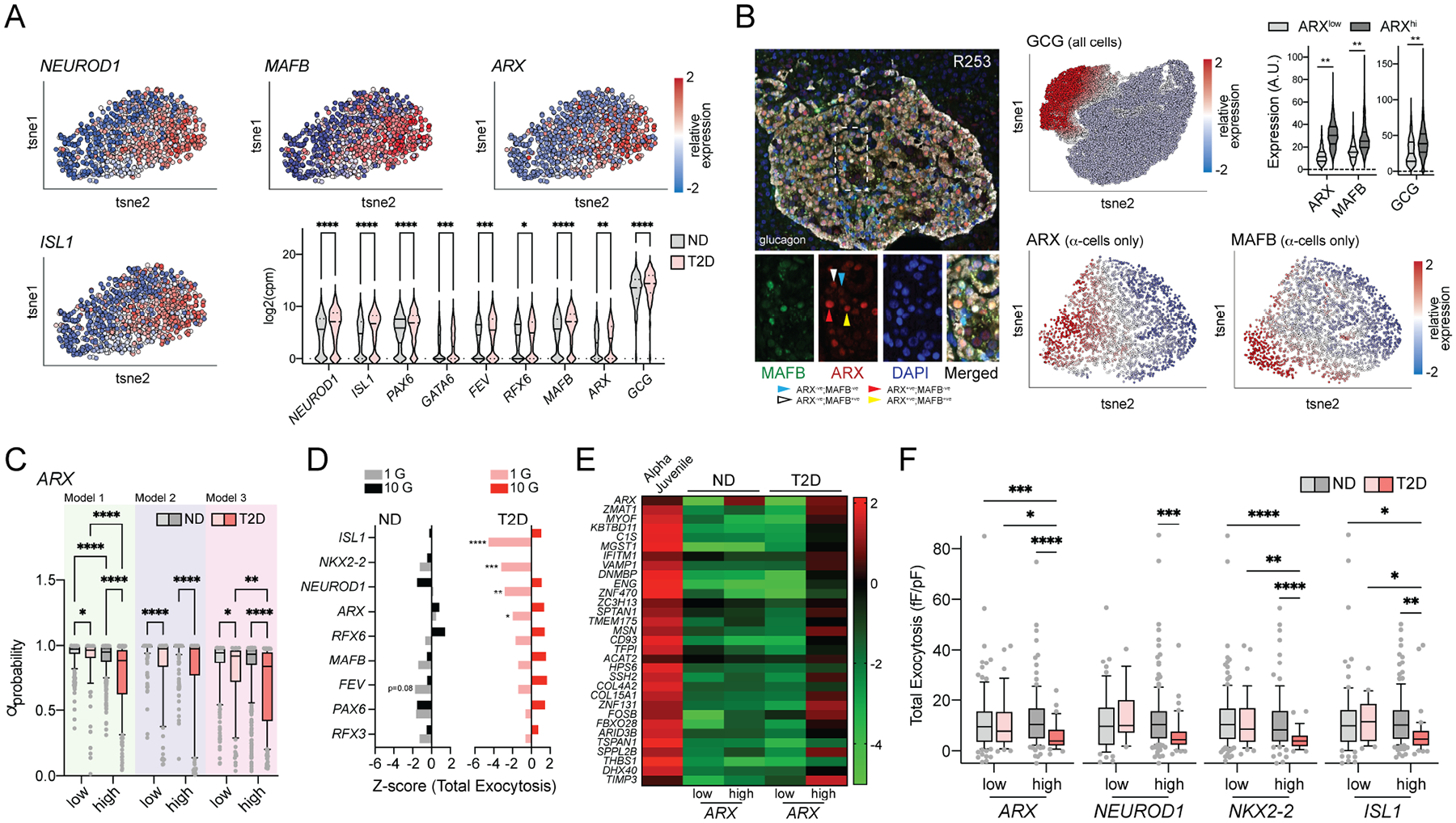
In diabetes, glucagon secretion from pancreatic α cells is dysregulated. The underlying mechanisms, and whether dysfunction occurs uniformly among cells, remain unclear. We examined α cells from human donors and mice using electrophysiological, transcriptomic, and computational approaches. Rising glucose suppresses α cell exocytosis by reducing P/Q-type Ca2+ channel activity, and this is disrupted in type 2 diabetes (T2D). Upon high-fat feeding of mice, α cells shift toward a "β cell-like" electrophysiological profile in concert with indications of impaired identity. In human α cells we identified links between cell membrane properties and cell surface signaling receptors, mitochondrial respiratory chain complex assembly, and cell maturation. Cell-type classification using machine learning of electrophysiology data demonstrated a heterogenous loss of "electrophysiologic identity" in α cells from donors with type 2 diabetes. Indeed, a subset of α cells with impaired exocytosis is defined by an enrichment in progenitor and lineage markers and upregulation of an immature transcriptomic phenotype, suggesting important links between α cell maturation state and dysfunction.
Keywords: alpha cells; diabetes; exocytosis; glucagon; human; islets of Langerhans; modeling; patch-seq.
Copyright © 2021 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Probing α-cell dysfunction in type 2 diabetes mellitus.Nat Rev Endocrinol. 2022 Apr;18(4):195. doi: 10.1038/s41574-022-00650-1. Nat Rev Endocrinol. 2022. PMID: 35210573 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
