

Natural triterpenoids from licorice potently inhibit SARS-CoV-2 infection
- PMID: 35116174
- PMCID: PMC8620242
- DOI: 10.1016/j.jare.2021.11.012
Natural triterpenoids from licorice potently inhibit SARS-CoV-2 infection
Abstract
Introduction: The COVID-19 global epidemic caused by severe acute respiratory syndrome coronavirus (SARS-CoV-2) is a great public health emergency. Discovering antiviral drug candidates is urgent for the prevention and treatment of COVID-19.
Objectives: This work aims to discover natural SARS-CoV-2 inhibitors from the traditional Chinese herbal medicine licorice.
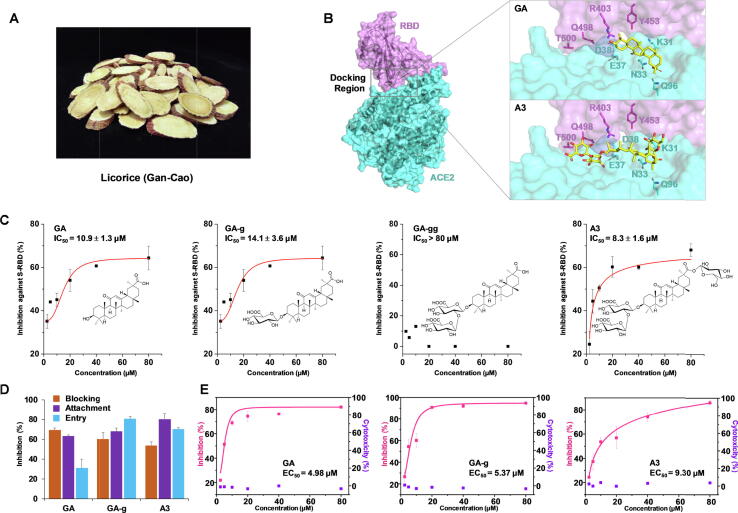
Methods: We screened 125 small molecules from Glycyrrhiza uralensis Fisch. (licorice, Gan-Cao) by virtual ligand screening targeting the receptor-binding domain (RBD) of SARS-CoV-2 spike protein. Potential hit compounds were further evaluated by ELISA, SPR, luciferase assay, antiviral assay and pharmacokinetic study.
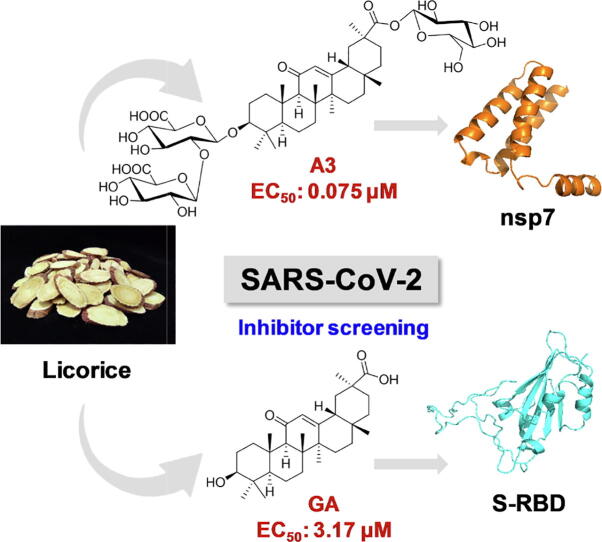
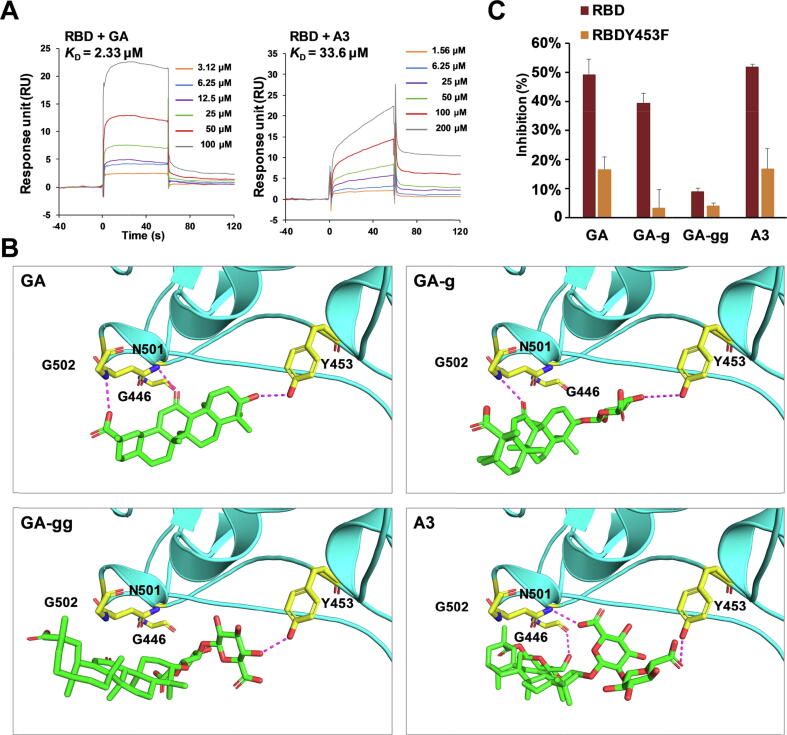
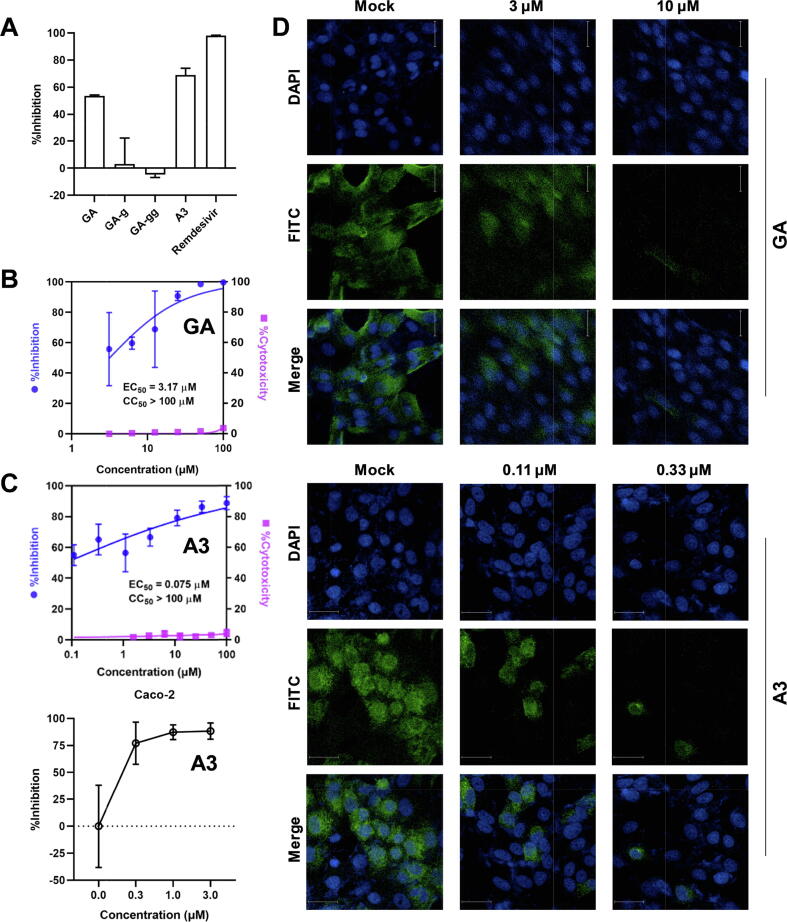
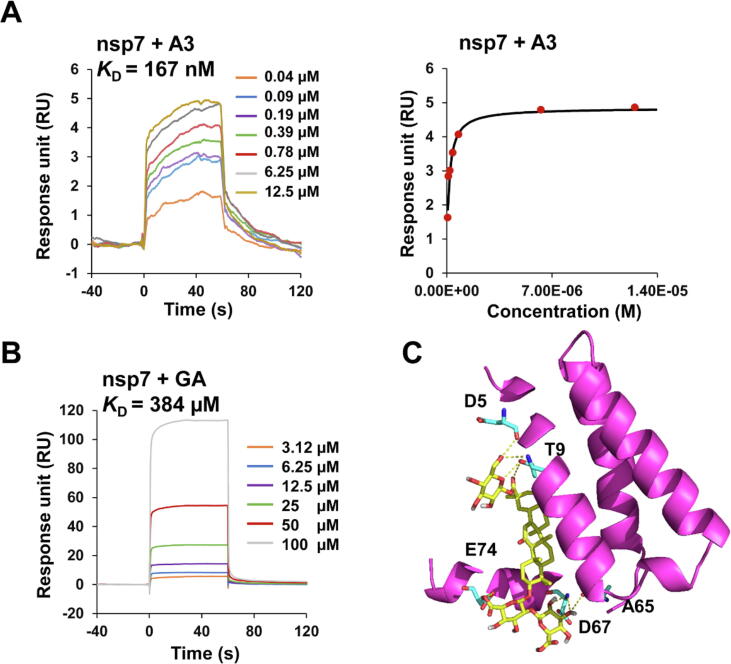
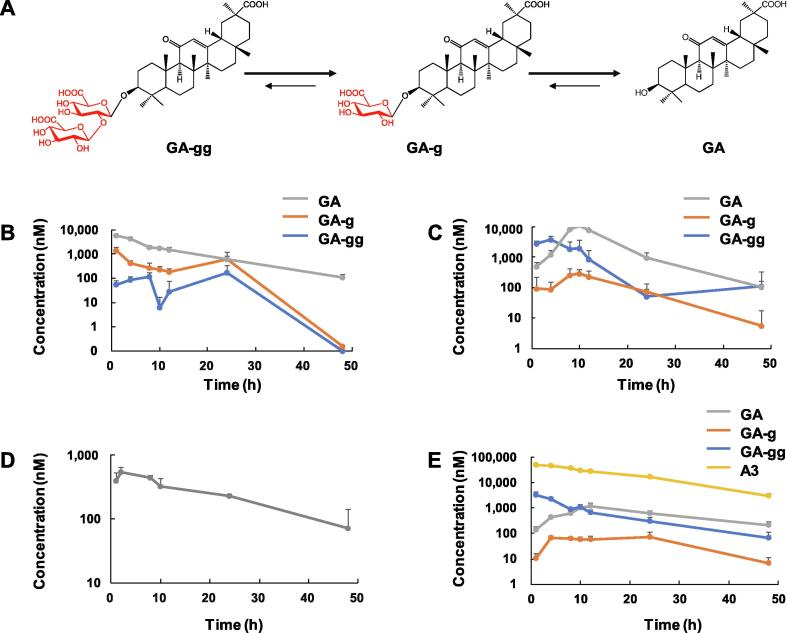
Results: The triterpenoids licorice-saponin A3 (A3) and glycyrrhetinic acid (GA) could potently inhibit SARS-CoV-2 infection, with EC50 of 75 nM and 3.17 µM, respectively. Moreover, we reveal that A3 mainly targets the nsp7 protein, and GA binds to the spike protein RBD of SARS-CoV-2.
Conclusion: In this work, we found GA and A3 from licorice potently inhibit SARS-CoV-2 infection by affecting entry and replication of the virus. Our findings indicate that these triterpenoids may contribute to the clinical efficacy of licorice for COVID-19 and could be promising candidates for antiviral drug development.
Keywords: COVID-19; Glycyrrhetinic acid; Licorice; Licorice-saponin A3; SARS-CoV-2.
© 2021 The Authors. Published by Elsevier B.V. on behalf of Cairo University.
Figures
References
-
- Cavanagh D. In: Coronaviruses with special emphasis on first insights concerning SARS. Schmidt A., Weber O., Wolff M.H., editors. Birkhauser Basel; Basel: 2005. Coronaviridae: a review of coronaviruses and toroviruses; pp. 1–54.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
