

Adipose-tissue plasticity in health and disease
- PMID: 35120662
- PMCID: PMC11152570
- DOI: 10.1016/j.cell.2021.12.016
Adipose-tissue plasticity in health and disease
Abstract
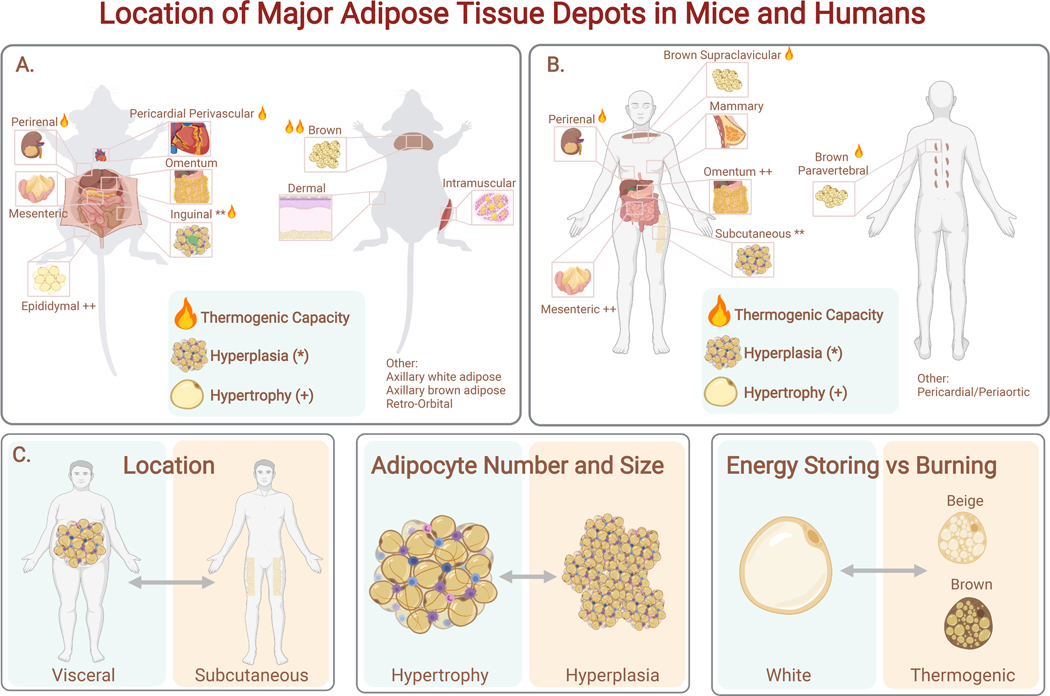
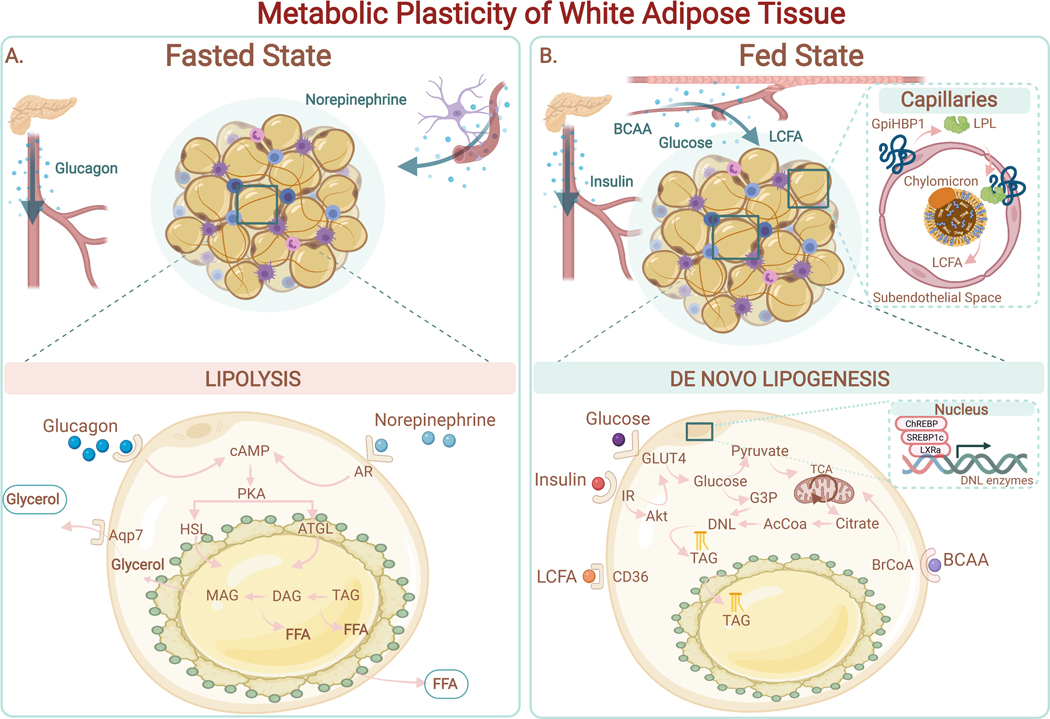
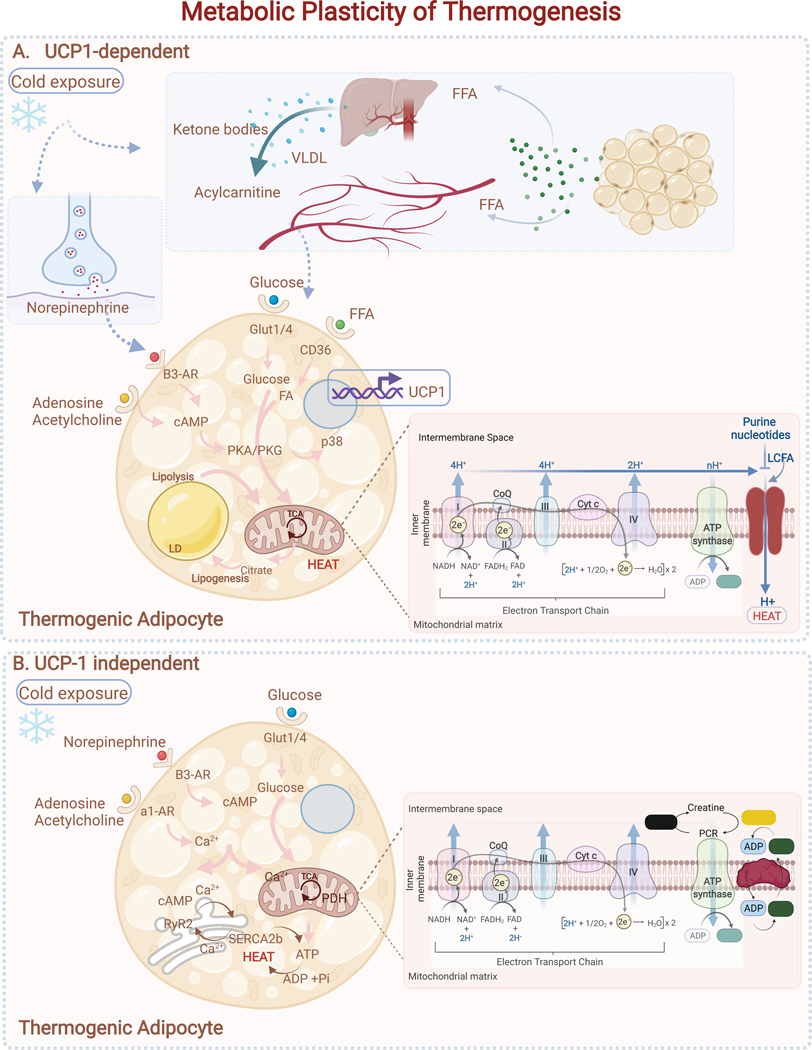
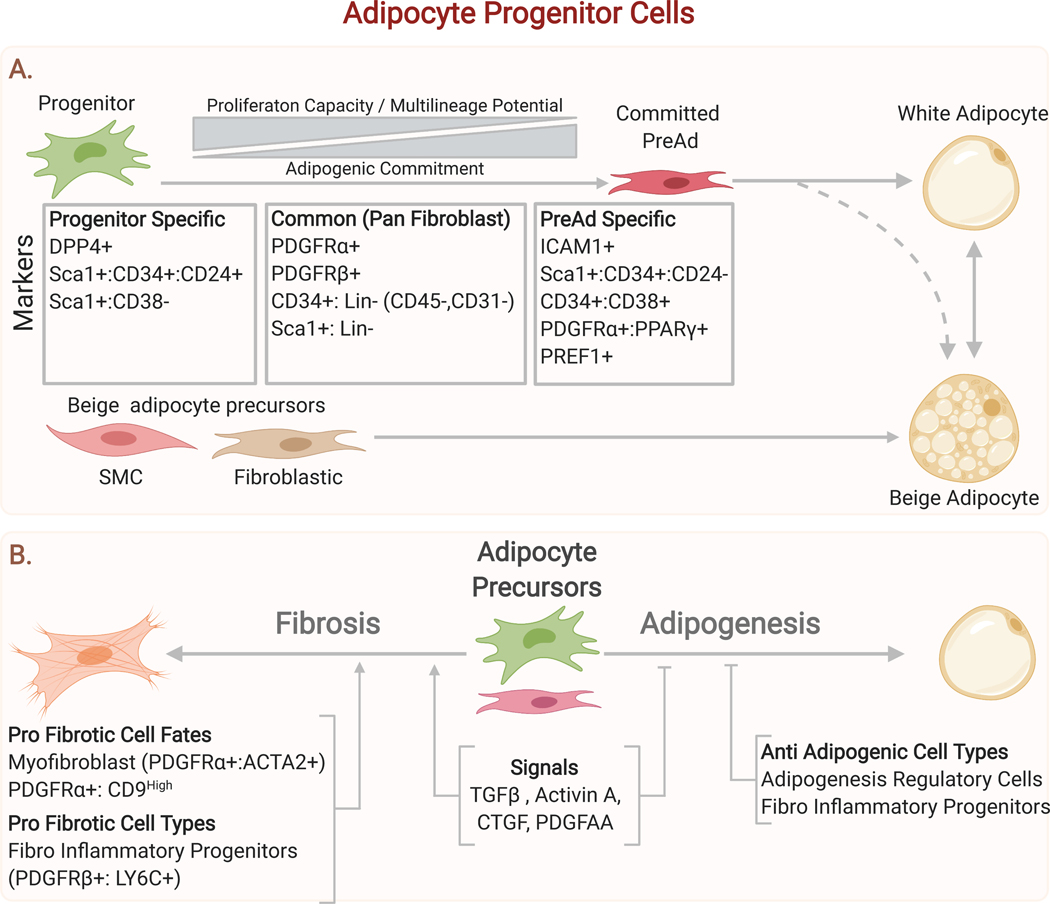
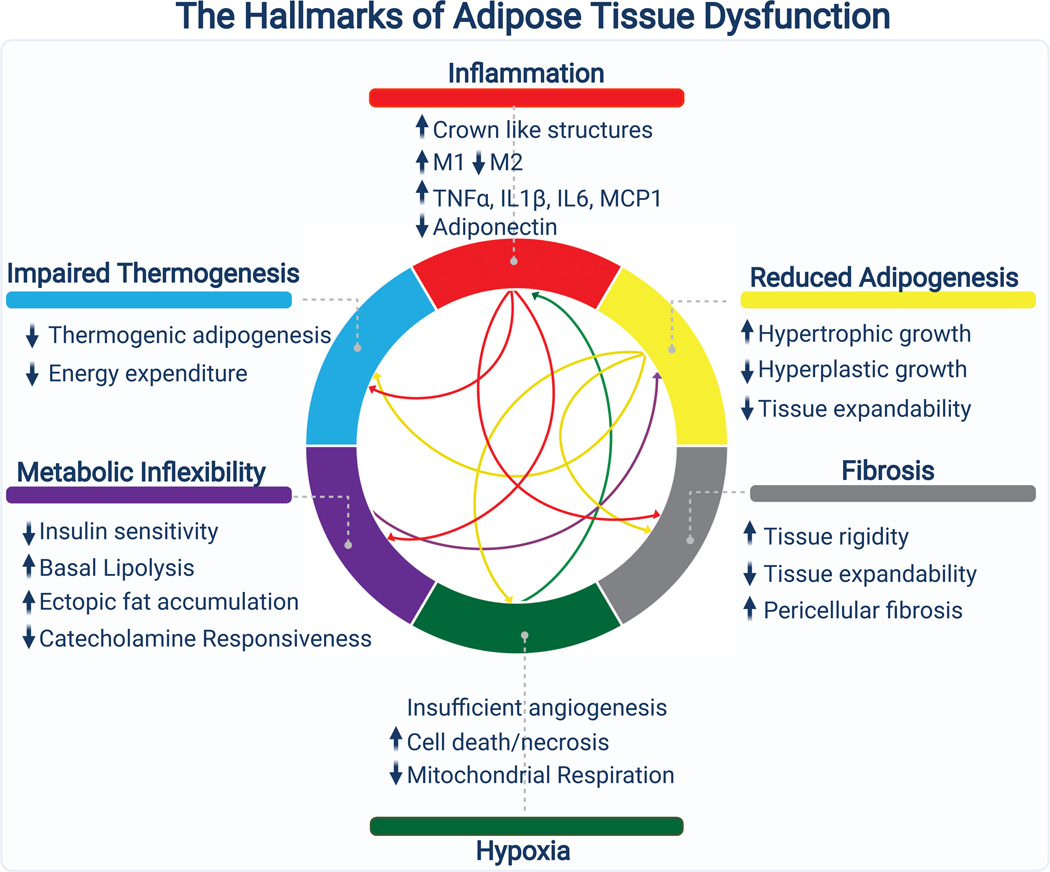
Adipose tissue, colloquially known as "fat," is an extraordinarily flexible and heterogeneous organ. While historically viewed as a passive site for energy storage, we now appreciate that adipose tissue regulates many aspects of whole-body physiology, including food intake, maintenance of energy levels, insulin sensitivity, body temperature, and immune responses. A crucial property of adipose tissue is its high degree of plasticity. Physiologic stimuli induce dramatic alterations in adipose-tissue metabolism, structure, and phenotype to meet the needs of the organism. Limitations to this plasticity cause diminished or aberrant responses to physiologic cues and drive the progression of cardiometabolic disease along with other pathological consequences of obesity.
Keywords: adipocyte; adipocyte progenitor; adipose tissue; beige fat; brown fat; diabetes; obesity; thermogenesis.
Copyright © 2021 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures

References
-
- Acosta JR, Douagi I, Andersson DP, Backdahl J, Ryden M, Arner P, and Laurencikiene J (2016). Increased fat cell size: a major phenotype of subcutaneous white adipose tissue in non-obese individuals with type 2 diabetes. Diabetologia 59, 560–570. - PubMed
-
- Andersson DP, Eriksson Hogling D, Thorell A, Toft E, Qvisth V, Naslund E, Thorne A, Wiren M, Lofgren P, Hoffstedt J, et al. (2014). Changes in subcutaneous fat cell volume and insulin sensitivity after weight loss. Diabetes Care 37, 1831–1836. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
