

Transmission potential of human schistosomes can be driven by resource competition among snail intermediate hosts
- PMID: 35121663
- PMCID: PMC8833218
- DOI: 10.1073/pnas.2116512119
Transmission potential of human schistosomes can be driven by resource competition among snail intermediate hosts
Abstract
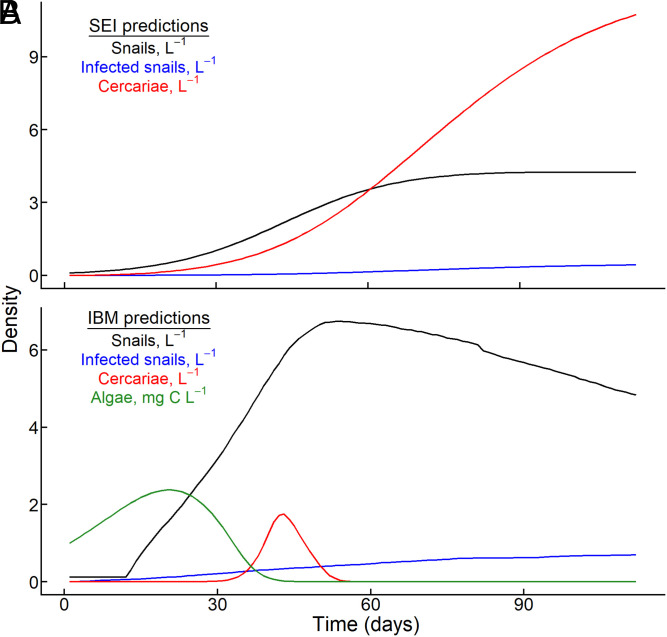
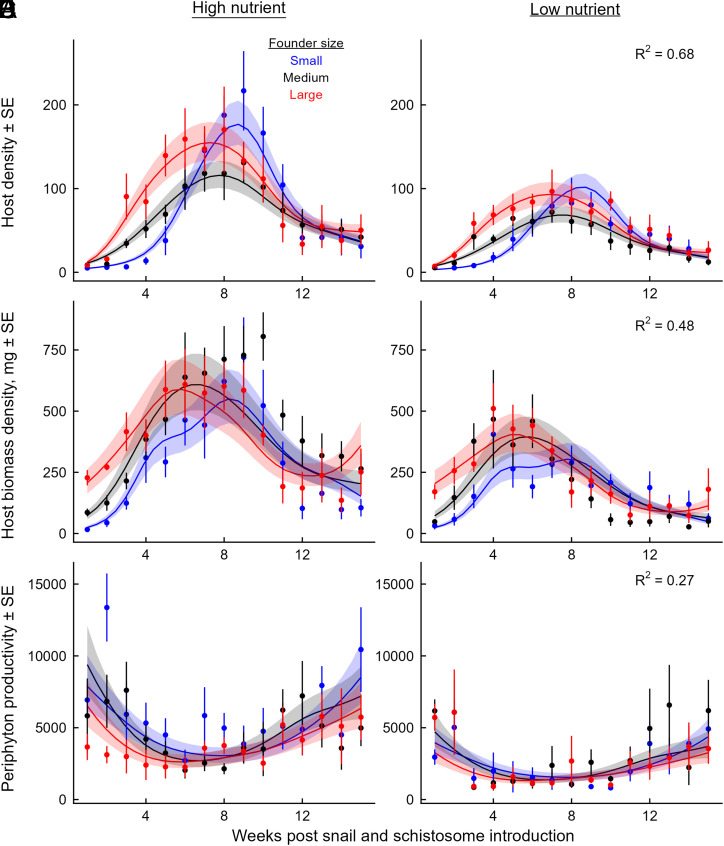
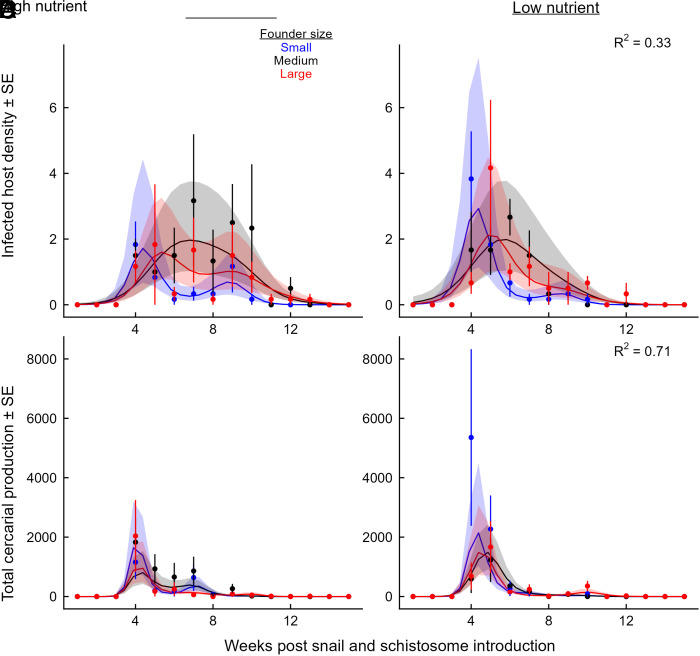
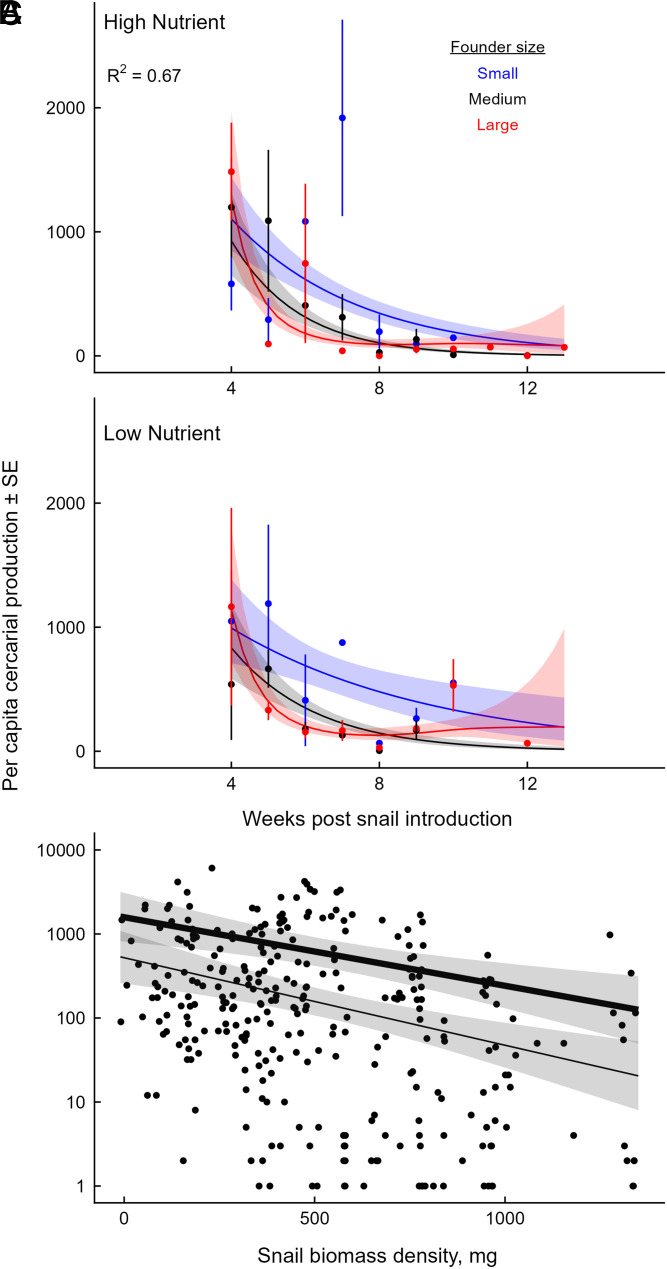
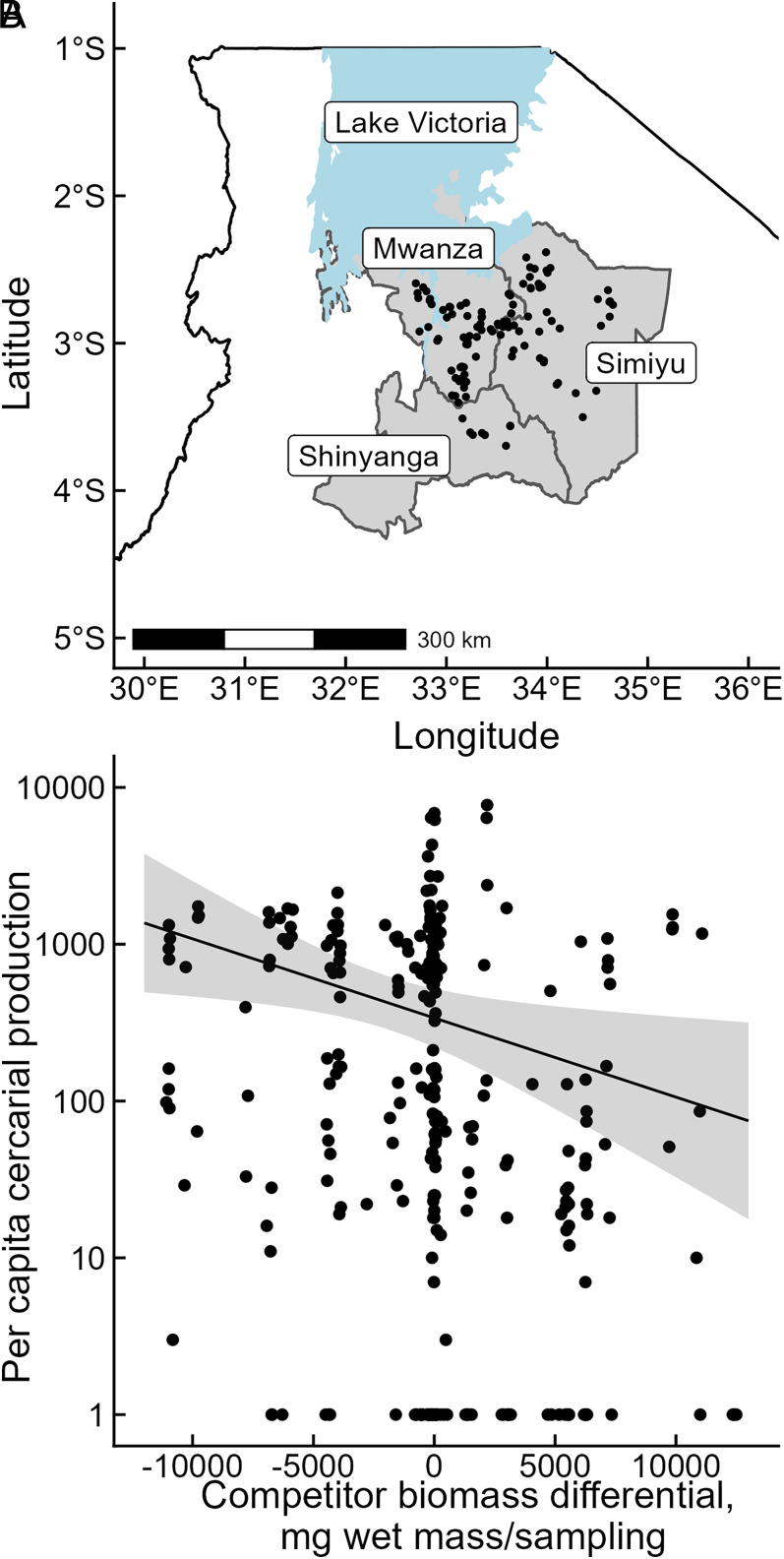
Predicting and disrupting transmission of human parasites from wildlife hosts or vectors remains challenging because ecological interactions can influence their epidemiological traits. Human schistosomes, parasitic flatworms that cycle between freshwater snails and humans, typify this challenge. Human exposure risk, given water contact, is driven by the production of free-living cercariae by snail populations. Conventional epidemiological models and management focus on the density of infected snails under the assumption that all snails are equally infectious. However, individual-level experiments contradict this assumption, showing increased production of schistosome cercariae with greater access to food resources. We built bioenergetics theory to predict how resource competition among snails drives the temporal dynamics of transmission potential to humans and tested these predictions with experimental epidemics and demonstrated consistency with field observations. This resource-explicit approach predicted an intense pulse of transmission potential when snail populations grow from low densities, i.e., when per capita access to resources is greatest, due to the resource-dependence of cercarial production. The experiment confirmed this prediction, identifying a strong effect of infected host size and the biomass of competitors on per capita cercarial production. A field survey of 109 waterbodies also found that per capita cercarial production decreased as competitor biomass increased. Further quantification of snail densities, sizes, cercarial production, and resources in diverse transmission sites is needed to assess the epidemiological importance of resource competition and support snail-based disruption of schistosome transmission. More broadly, this work illustrates how resource competition can sever the correspondence between infectious host density and transmission potential.
Keywords: energy budget; parasitism; resource competition; schistosome; transmission potential.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Dijkstra K. B., Schrama M. J. J., Gorsich E. E., Hochkirch A., “Deadly mosquito” or “living freshwater”? Science 361, 341 (2018). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
