

The SINGLE FLOWER (SFL) gene encodes a MYB transcription factor that regulates the number of flowers produced by the inflorescence of chickpea
- PMID: 35122280
- PMCID: PMC9314632
- DOI: 10.1111/nph.18019
The SINGLE FLOWER (SFL) gene encodes a MYB transcription factor that regulates the number of flowers produced by the inflorescence of chickpea
Abstract
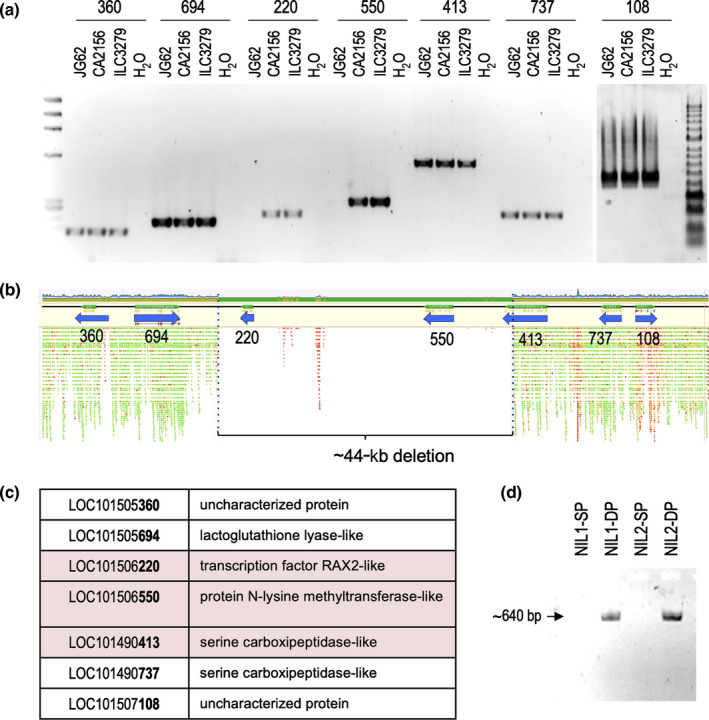
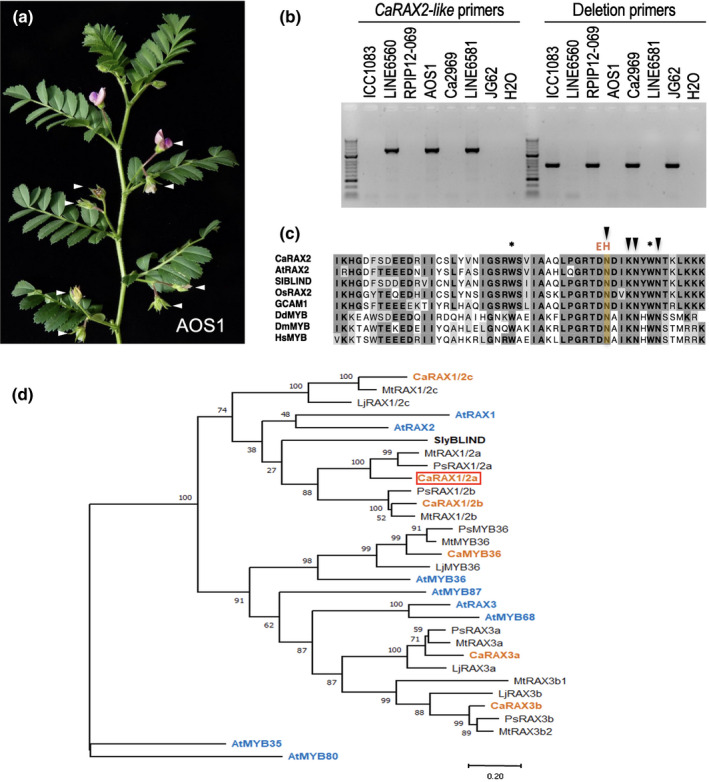
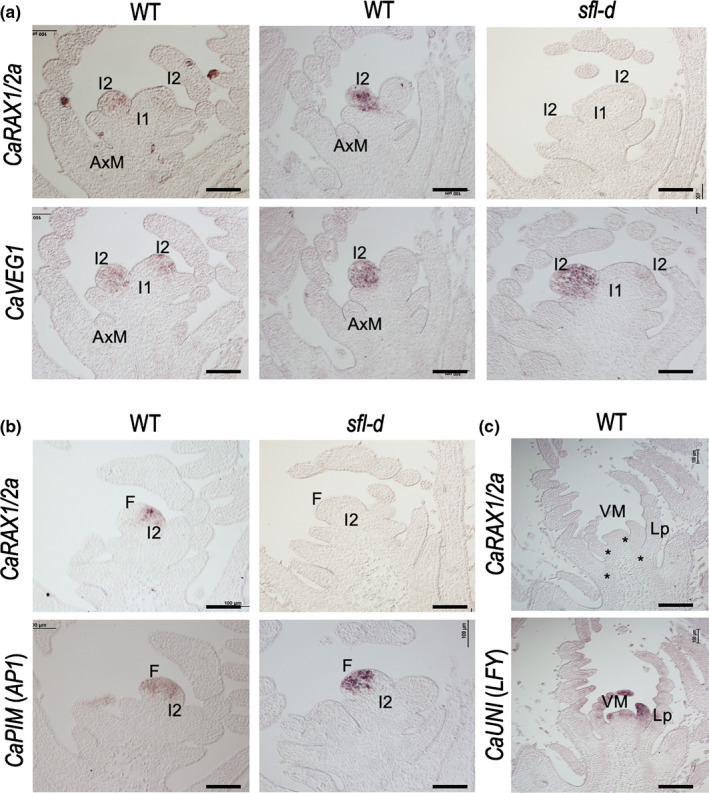
Legumes usually have compound inflorescences, where flowers/pods develop from secondary inflorescences (I2), formed laterally at the primary inflorescence (I1). Number of flowers per I2, characteristic of each legume species, has important ecological and evolutionary relevance as it determines diversity in inflorescence architecture; moreover, it is also agronomically important for its potential impact on yield. Nevertheless, the genetic network controlling the number of flowers per I2 is virtually unknown. Chickpea (Cicer arietinum) typically produces one flower per I2 but single flower (sfl) mutants produce two (double-pod phenotype). We isolated the SFL gene by mapping the sfl-d mutation and identifying and characterising a second mutant allele. We analysed the effect of sfl on chickpea inflorescence ontogeny with scanning electron microscopy and studied the expression of SFL and meristem identity genes by RNA in situ hybridisation. We show that SFL corresponds to CaRAX1/2a, which codes a MYB transcription factor specifically expressed in the I2 meristem. Our findings reveal SFL as a central factor controlling chickpea inflorescence architecture, acting in the I2 meristem to regulate the length of the period for which it remains active, and therefore determining the number of floral meristems that it can produce.
Keywords: R2R3-MYB genes; RAX genes; chickpea breeding; chickpea double-pod mutants; compound inflorescence; inflorescence architecture; meristem activity; seed yield.
© 2022 The Authors. New Phytologist © 2022 New Phytologist Foundation.
Figures

References
-
- Ali L, Deokar A, Caballo C, Tar’an B, Gil J, Chen W, Millan T, Rubio J. 2016. Fine mapping for double podding gene in chickpea. Theoretical and Applied Genetics 129: 77–86. - PubMed
-
- Berbel A, Ferrándiz C, Hecht V, Dalmais M, Lund OS, Sussmilch FC, Taylor SA, Bendahmane A, Ellis THN, Beltrán JP et al. 2012. VEGETATIVE1 is essential for development of the compound inflorescence in pea. Nature Communications 3: 659–714. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
