

FNDC5/irisin facilitates muscle-adipose-bone connectivity through ubiquitination-dependent activation of runt-related transcriptional factors RUNX1/2
- PMID: 35124008
- PMCID: PMC8892030
- DOI: 10.1016/j.jbc.2022.101679
FNDC5/irisin facilitates muscle-adipose-bone connectivity through ubiquitination-dependent activation of runt-related transcriptional factors RUNX1/2
Abstract
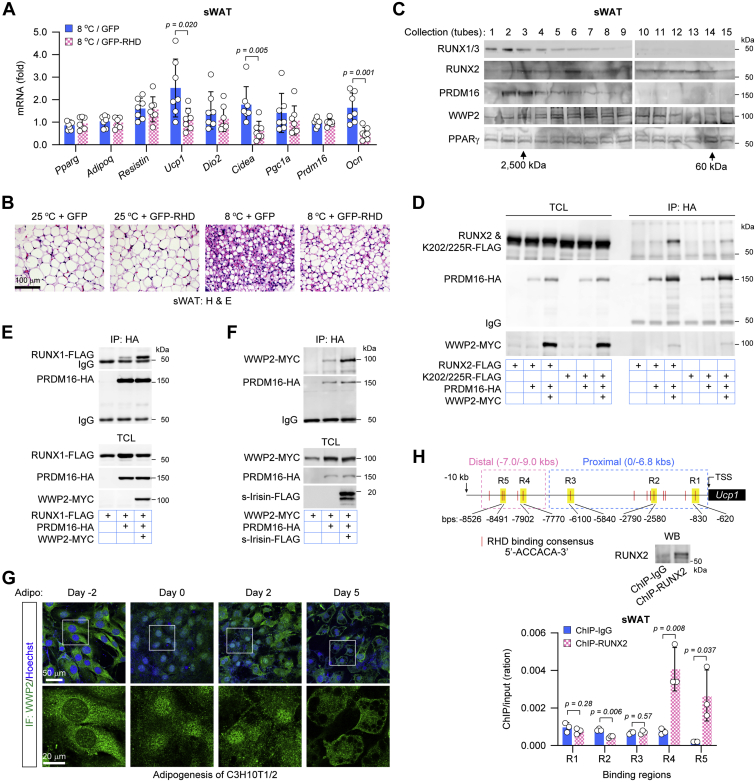
In the past decade, the cleavage protein irisin derived from fibronectin type III domain-containing protein 5 (FNDC5) in exercise-stimulated skeletal muscle has increasingly become a biomarker associated with metabolic syndrome and osteoporosis in humans. However, it is unclear how this protein facilitates muscle-adipose-bone connectivity in metabolic and skeletal homeostasis. In this study, we unexpectedly observed that the FNDC5 gene can be markedly activated during the differentiation of brown adipocytes but not white adipocytes, and that FNDC5 is specifically expressed in mouse brown adipose tissues (BATs). But unlike it in the skeletal muscles, the expression of FNDC5/irisin in BAT is promoted by cold exposure rather than exercise in mice. Analysis of promoter activity and chromatin immunoprecipitation further showed that peroxisome proliferator-activated receptor γ coactivator-1α and thyroid hormone receptors cooperate on the FNDC5 gene promoter to induce its transcription. We found that FNDC5/irisin stimulates the runt-related transcriptional factors RUNX1/2 via a focal adhesion kinase-dependent pathway in both bone and subcutaneous white adipose tissues. Mechanistically, focal adhesion kinase is stimulated by FNDC5/irisin and then facilitates E3 ubiquitin-protein ligase WW domain-containing protein 2 to ubiquitinate and subsequently activate RUNX1/2, culminating in the activation of osteoblast-related or thermogenesis-related genes. Interestingly, the PR domain containing protein 16 that is crucial for subcutaneous white adipose "browning" and skeletal development was found to form a complex with RUNX1/2 in a WW domain-containing protein 2-dependent manner. These findings elucidate a signaling mechanism by which FNDC5/irisin supports the muscle-adipose-bone connectivity, especially BAT-bone connectivity.
Keywords: WW domain–containing protein 2; brown adipose tissue; fibronectin type III domain–containing protein 5/irisin; focal adhesion kinase; the runt-related transcriptional factor 1/2; white adipose tissue.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures






References
-
- Virtanen K.A., Lidell M.E., Orava J., Heglind M., Westergren R., Niemi T., Taittonen M., Laine J., Savisto N.J., Enerbäck S., Nuutila P. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 2009;360:1518–1525. - PubMed
-
- Enerback S. Human brown adipose tissue. Cell Metab. 2010;11:248–252. - PubMed
-
- Wu J., Boström P., Sparks L.M., Ye L., Choi J.H., Giang A.H., Khandekar M., Virtanen K.A., Nuutila P., Schaart G., Huang K., Tu H., van Marken Lichtenbelt W.D., Hoeks J., Enerbäck S., et al. Beige adipocytes are a distinct type of thermogenic fat cell in mouse and human. Cell. 2012;150:66–376. - PMC - PubMed
-
- Himms-Hagen J., Melnyk A., Zingaretti M.C., Ceresi E., Barbatelli G., Cinti S. Multilocular fat cells in WAT of CL-316243-treated rats derive directly from white adipocytes. Am. J. Physiol. Cell Physiol. 2000;279:C670–C681. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
