

Regulation of the activity of the bacterial histidine kinase PleC by the scaffolding protein PodJ
- PMID: 35124010
- PMCID: PMC8980812
- DOI: 10.1016/j.jbc.2022.101683
Regulation of the activity of the bacterial histidine kinase PleC by the scaffolding protein PodJ
Abstract
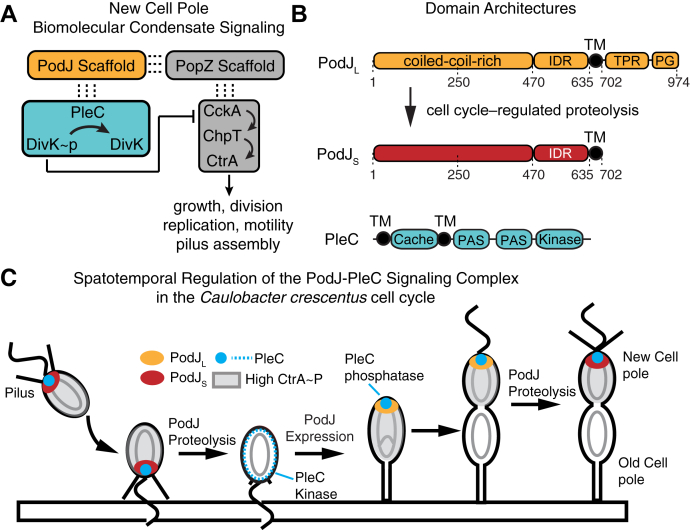
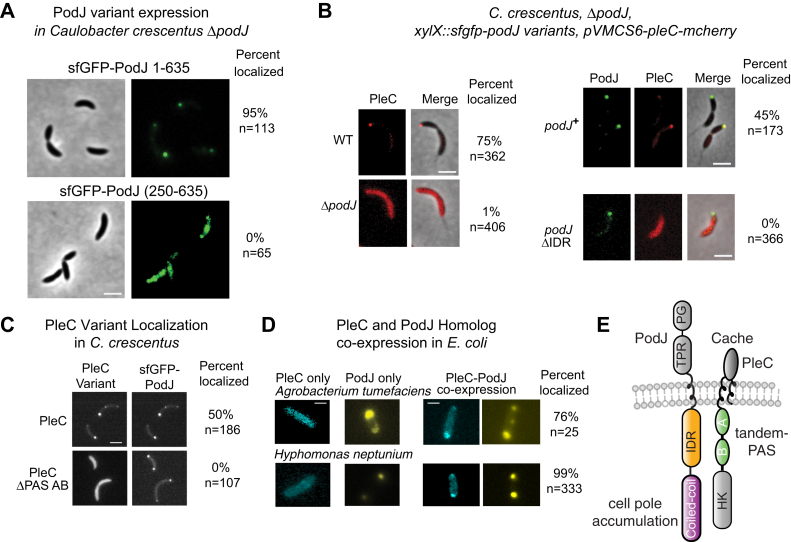
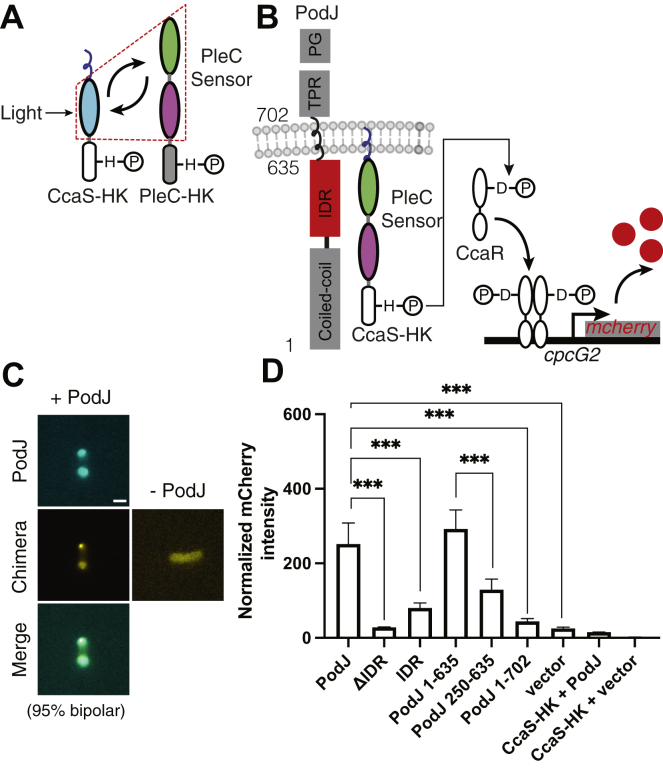
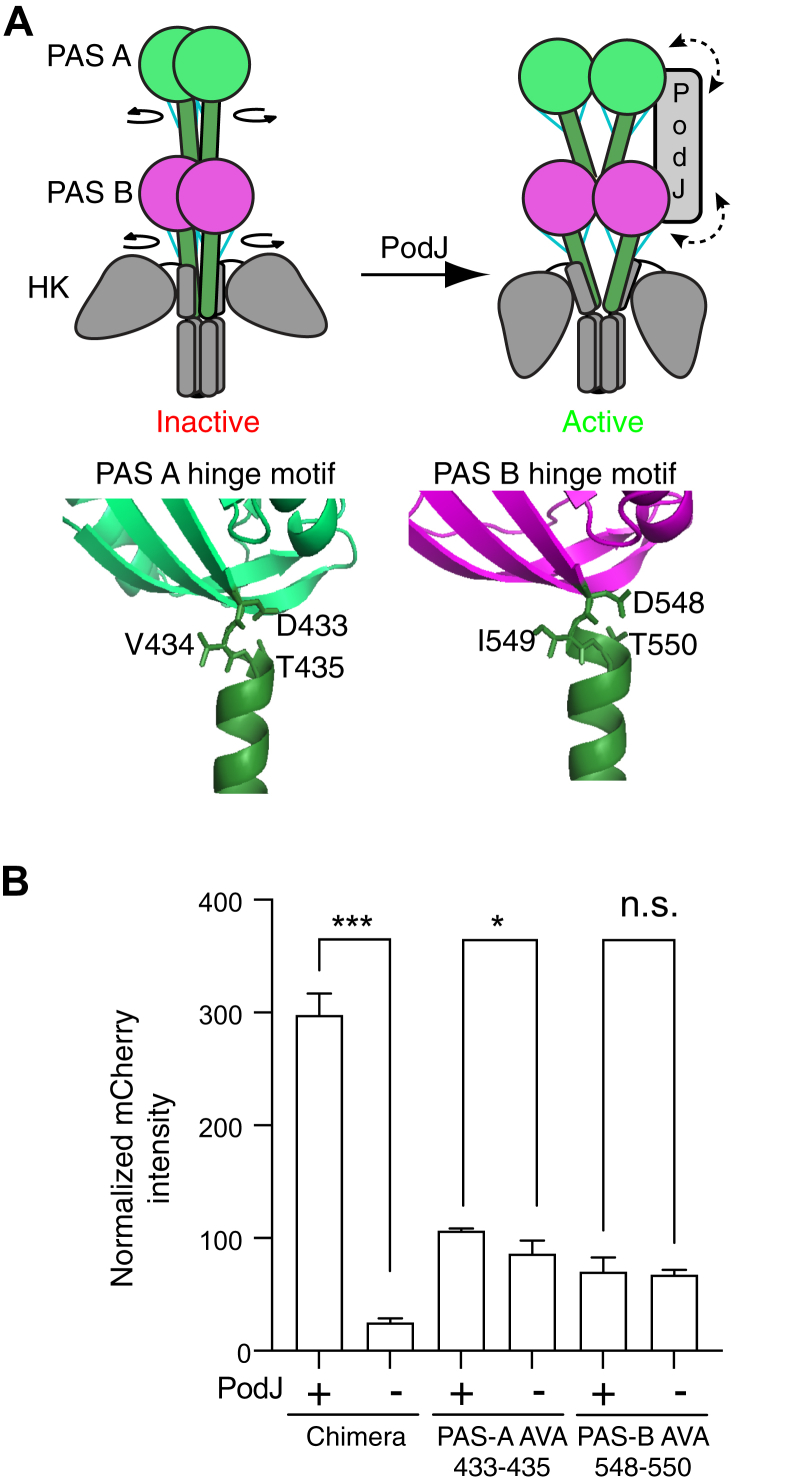
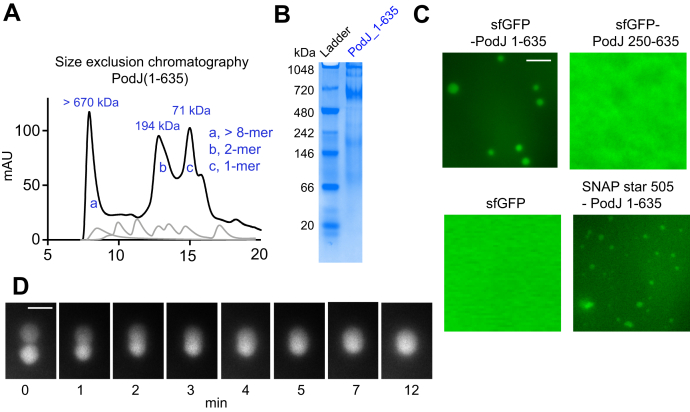
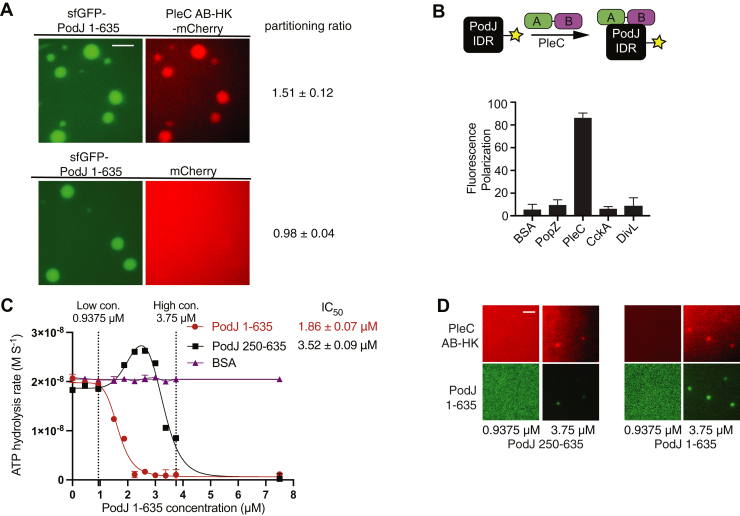
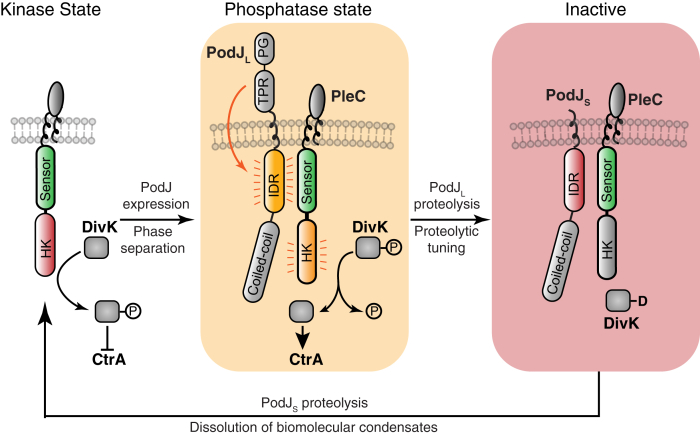
Scaffolding proteins can customize the response of signaling networks to support cell development and behaviors. PleC is a bifunctional histidine kinase whose signaling activity coordinates asymmetric cell division to yield a motile swarmer cell and a stalked cell in the gram-negative bacterium Caulobacter crescentus. Past studies have shown that PleC's switch in activity from kinase to phosphatase correlates with a change in its subcellular localization pattern from diffuse to localized at the new cell pole. Here we investigated how the bacterial scaffolding protein PodJ regulates the subcellular positioning and activity of PleC. We reconstituted the PleC-PodJ signaling complex through both heterologous expressions in Escherichia coli and in vitro studies. In vitro, PodJ phase separates as a biomolecular condensate that recruits PleC and inhibits its kinase activity. We also constructed an in vivo PleC-CcaS chimeric histidine kinase reporter assay and demonstrated using this method that PodJ leverages its intrinsically disordered region to bind to PleC's PAS sensory domain and regulate PleC-CcaS signaling. Regulation of the PleC-CcaS was most robust when PodJ was concentrated at the cell poles and was dependent on the allosteric coupling between PleC-CcaS's PAS sensory domain and its downstream histidine kinase domain. In conclusion, our in vitro biochemical studies suggest that PodJ phase separation may be coupled to changes in PleC enzymatic function. We propose that this coupling of phase separation and allosteric regulation may be a generalizable phenomenon among enzymes associated with biomolecular condensates.
Keywords: Caulobacter crescentus; PAS domain; biomolecular condensate; histidine kinase; intrinsically disordered region (IDR); phase separation; scaffold.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
-
- Bhattacharyya R.P., Remenyi A., Good M.C., Bashor C.J., Falick A.M., Lim W.A. The Ste5 scaffold allosterically modulates signaling output of the yeast mating pathway. Science. 2006;311:822–826. - PubMed
-
- Cohan M.C., Pappu R.V. Making the case for disordered proteins and biomolecular condensates in bacteria. Trends Biochem. Sci. 2020;45:668–680. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
