

Genome-wide evolutionary response of European oaks during the Anthropocene
- PMID: 35127134
- PMCID: PMC8802238
- DOI: 10.1002/evl3.269
Genome-wide evolutionary response of European oaks during the Anthropocene
Abstract
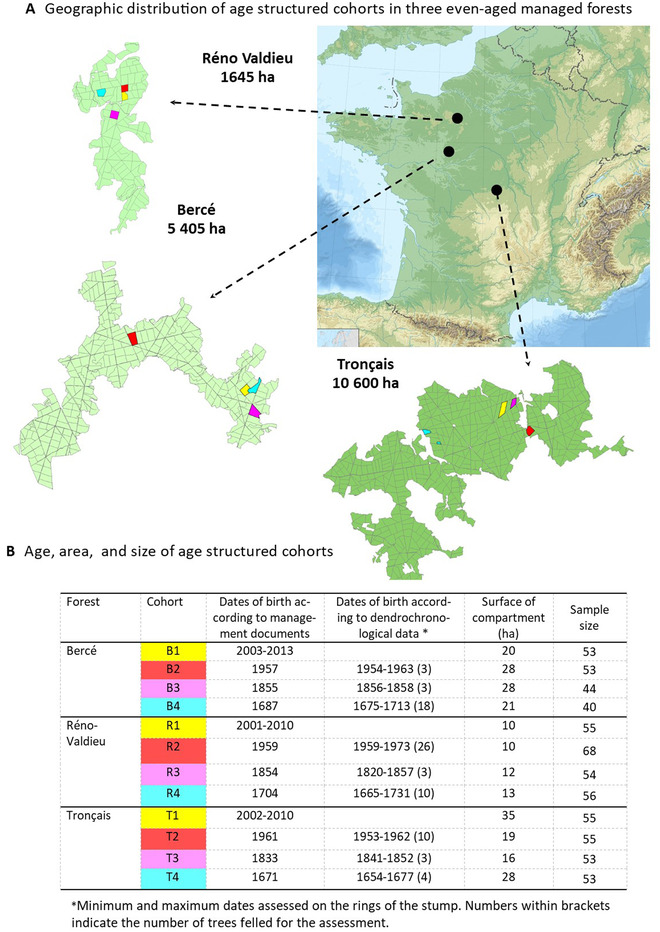
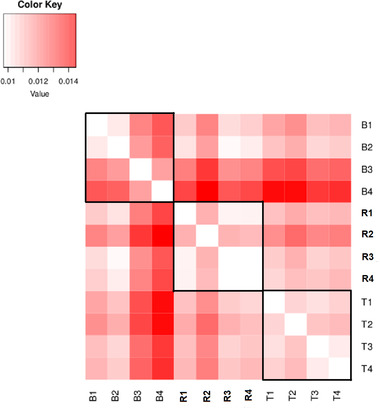
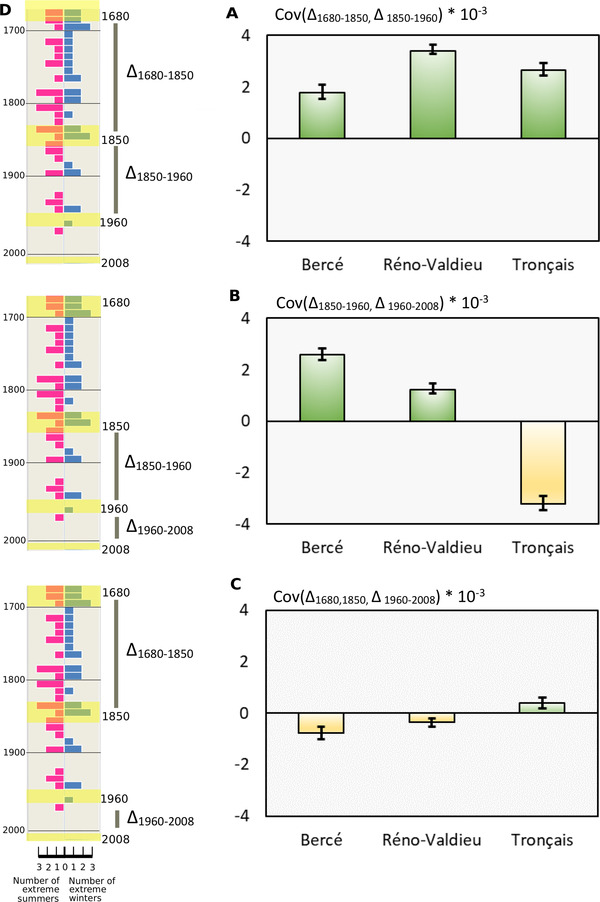
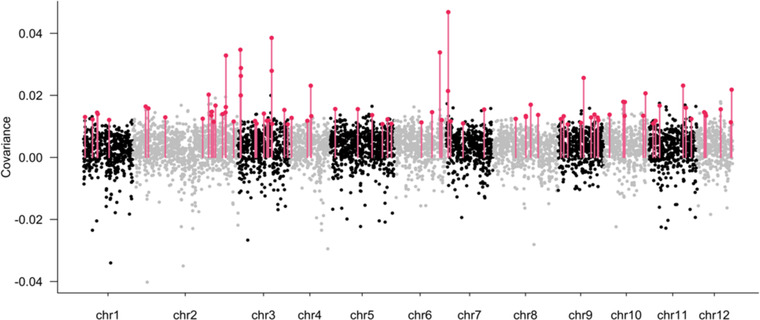
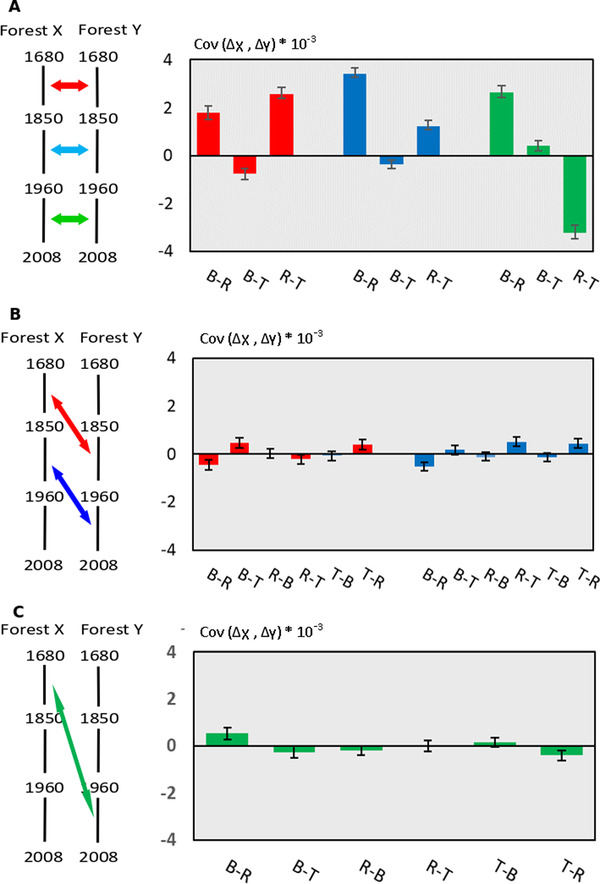
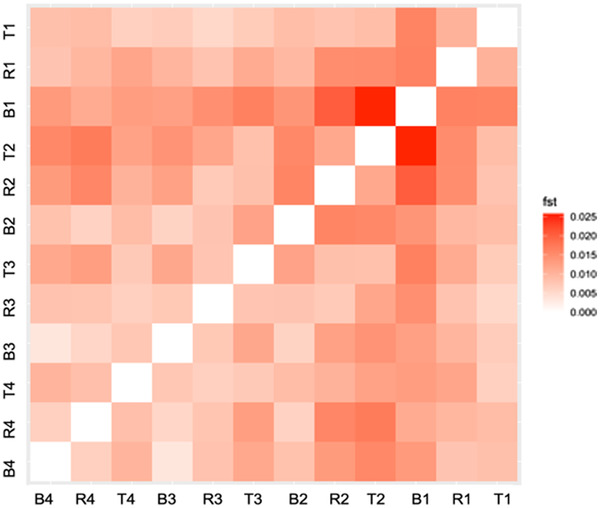
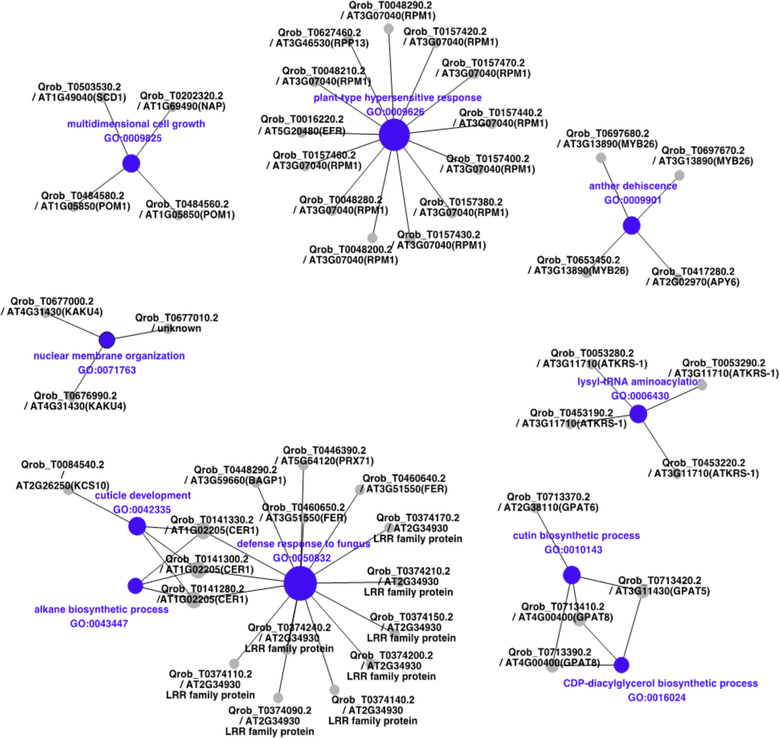
The pace of tree microevolution during Anthropocene warming is largely unknown. We used a retrospective approach to monitor genomic changes in oak trees since the Little Ice Age (LIA). Allelic frequency changes were assessed from whole-genome pooled sequences for four age-structured cohorts of sessile oak (Quercus petraea) dating back to 1680, in each of three different oak forests in France. The genetic covariances of allelic frequency changes increased between successive time periods, highlighting genome-wide effects of linked selection. We found imprints of parallel linked selection in the three forests during the late LIA, and a shift of selection during more recent time periods of the Anthropocene. The changes in allelic covariances within and between forests mirrored the documented changes in the occurrence of extreme events (droughts and frosts) over the last 300 years. The genomic regions with the highest covariances were enriched in genes involved in plant responses to pathogens and abiotic stresses (temperature and drought). These responses are consistent with the reported sequence of frost (or drought) and disease damage ultimately leading to the oak dieback after extreme events. They provide support for adaptive evolution of long-lived species during recent climatic changes. Although we acknowledge that other sources (e.g., gene flow, generation overlap) may have contributed to temporal covariances of allelic frequency changes, the consistent and correlated response across the three forests lends support to the existence of a systematic driving force such as natural selection.
Keywords: Anthropocene; Little Ice Age; Quercus petraea; evolution; linked selection.
© 2021 The Authors. Evolution Letters published by Wiley Periodicals LLC on behalf of Society for the Study of Evolution (SSE) and European Society for Evolutionary Biology (ESEB).
Figures
References
-
- Alberto, F. , Bouffier L., Louvet J. M., Lamy J. B., Delzon S., and Kremer A.. 2011. Adaptive responses for seed and leaf phenology in natural populations of sessile oak along an altitudinal gradient. J. Evol. Biol. 24:1442–1454. - PubMed
-
- Alexa, A. , and Rahnenfuhrer J.. 2020. topGO: enrichment analysis for gene ontology. R package version 2.40.0. Available via https://bioconductor.org/packages/release/bioc/html/topGO.html.
-
- Alexa, A. , Rahnenfuhrer J., and Lengauer T.. 2006. Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 22:1600–1607. - PubMed
-
- Altmann, A. , Weber P., Bader D., Preuss M., Binder E. B., and Muller‐Myhsok B.. 2012. A beginners guide to SNP calling from high‐throughput DNA‐sequencing data. Hum. Genet. 131:1541–1554. - PubMed
