

Sex differences in endurance exercise capacity and skeletal muscle lipid metabolism in mice
- PMID: 35133078
- PMCID: PMC8822869
- DOI: 10.14814/phy2.15174
Sex differences in endurance exercise capacity and skeletal muscle lipid metabolism in mice
Abstract
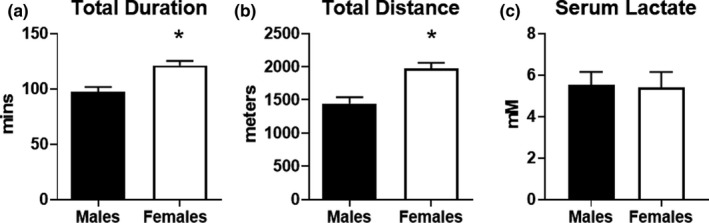
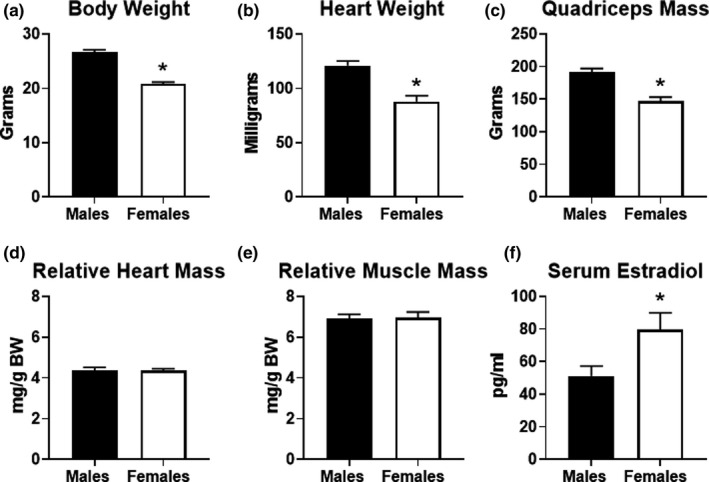
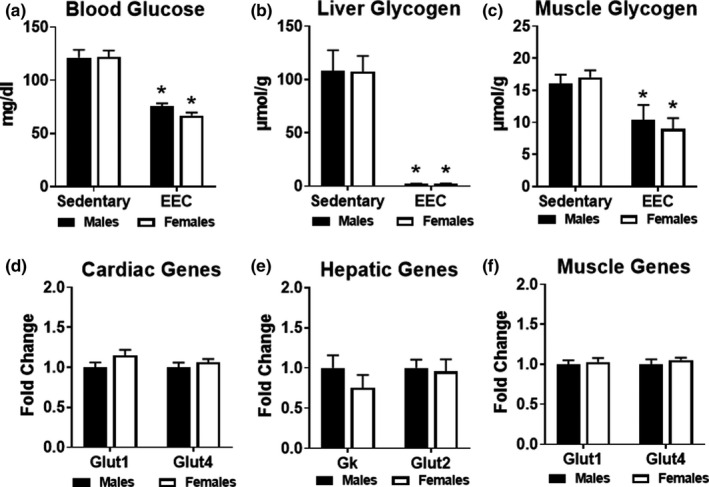
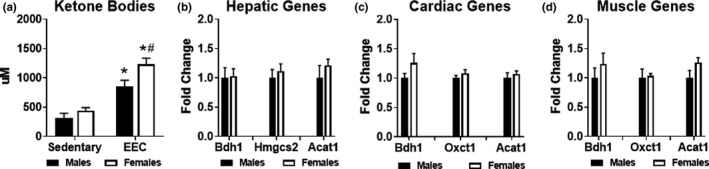
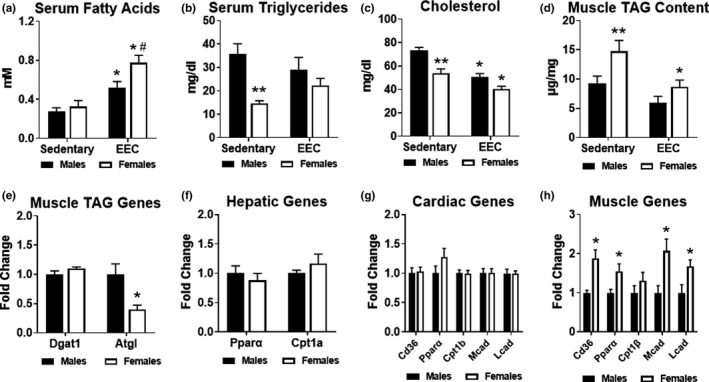
Previous studies suggest that sex differences in lipid metabolism exist with females demonstrating a higher utilization of lipids during exercise, which is mediated partly by increased utilization of muscle triglycerides. However, whether these changes in lipid metabolism contribute directly to endurance exercise performance is unclear. Therefore, the objective of this study was to investigate the contribution of exercise substrate metabolism to sex differences in endurance exercise capacity (EEC) in mice. Male and female C57BL/6-NCrl mice were subjected to an EEC test until exhaustion on a motorized treadmill. The treadmill was set at a 10% incline, and the speed gradually increased from 10.2 m/min to 22.2 m/min at fixed intervals for up to 2.5 h. Tissues and blood were harvested in mice immediately following the EEC. A cohort of sedentary, non-exercised male and female mice were used as controls. Females outperformed males by ~25% on the EEC. Serum levels of both fatty acids and ketone bodies were ~50% higher in females at the end of the EEC. In sedentary female mice, skeletal muscle triglyceride content was significantly greater compared to sedentary males. Gene expression analysis demonstrated that genes involved in skeletal muscle fatty acid oxidation were significantly higher in females with no changes in genes associated with glucose uptake or ketone body oxidation. The findings suggest that female mice have a higher endurance exercise capacity and a greater ability to mobilize and utilize fatty acids for energy.
Keywords: exercise metabolism; exercise physiology; fatty acid oxidation; ketosis; triglyceride metabolism.
© 2022 The Authors. Physiological Reports published by Wiley Periodicals LLC on behalf of The Physiological Society and the American Physiological Society.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
Similar articles
-
L-Carnitine enhances exercise endurance capacity by promoting muscle oxidative metabolism in mice.Biochem Biophys Res Commun. 2015 Aug 21;464(2):568-73. doi: 10.1016/j.bbrc.2015.07.009. Epub 2015 Jul 8. Biochem Biophys Res Commun. 2015. PMID: 26164228
-
Green tea extract improves running endurance in mice by stimulating lipid utilization during exercise.Am J Physiol Regul Integr Comp Physiol. 2006 Jun;290(6):R1550-6. doi: 10.1152/ajpregu.00752.2005. Epub 2006 Jan 12. Am J Physiol Regul Integr Comp Physiol. 2006. PMID: 16410398
-
Effects of short-term exercise and endurance training on skeletal muscle mitochondria damage induced by particular matter, atmospherically relevant artificial PM2.5.Front Public Health. 2024 Feb 28;12:1302175. doi: 10.3389/fpubh.2024.1302175. eCollection 2024. Front Public Health. 2024. PMID: 38481847 Free PMC article.
-
Triglyceride metabolism in exercising muscle.Biochim Biophys Acta Mol Cell Biol Lipids. 2017 Oct;1862(10 Pt B):1250-1259. doi: 10.1016/j.bbalip.2017.06.015. Epub 2017 Jun 24. Biochim Biophys Acta Mol Cell Biol Lipids. 2017. PMID: 28652193 Review.
-
An "Exercise" in Cardiac Metabolism.Front Cardiovasc Med. 2018 Jun 7;5:66. doi: 10.3389/fcvm.2018.00066. eCollection 2018. Front Cardiovasc Med. 2018. PMID: 29930946 Free PMC article. Review.
Cited by
-
Reveal key genes and factors affecting athletes performance in endurance sports using bioinformatic technologies.BMC Genom Data. 2023 Feb 21;24(1):10. doi: 10.1186/s12863-023-01106-9. BMC Genom Data. 2023. PMID: 36809973 Free PMC article.
-
Age-related changes of skeletal muscle metabolic response to contraction are also sex-dependent.J Physiol. 2025 Jan;603(1):69-86. doi: 10.1113/JP285124. Epub 2023 Sep 23. J Physiol. 2025. PMID: 37742081 Free PMC article.
-
Effects of defined voluntary running distances coupled with high-fat diet consumption on the skeletal muscle transcriptome of male mice.Physiol Rep. 2025 Jan;13(2):e70170. doi: 10.14814/phy2.70170. Physiol Rep. 2025. PMID: 39821584 Free PMC article.
-
SMURF1-Induced Ubiquitination of FTH1 Disrupts Iron Homeostasis and Suppresses Myogenesis.Int J Mol Sci. 2025 Feb 6;26(3):1390. doi: 10.3390/ijms26031390. Int J Mol Sci. 2025. PMID: 39941157 Free PMC article.
-
The influence of sex on activity in voluntary wheel running, forced treadmill running, and open field testing in mice.Physiol Rep. 2025 Feb;13(4):e70246. doi: 10.14814/phy2.70246. Physiol Rep. 2025. PMID: 39985141 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
