

Physiological synaptic activity and recognition memory require astroglial glutamine
- PMID: 35136061
- PMCID: PMC8826940
- DOI: 10.1038/s41467-022-28331-7
Physiological synaptic activity and recognition memory require astroglial glutamine
Abstract
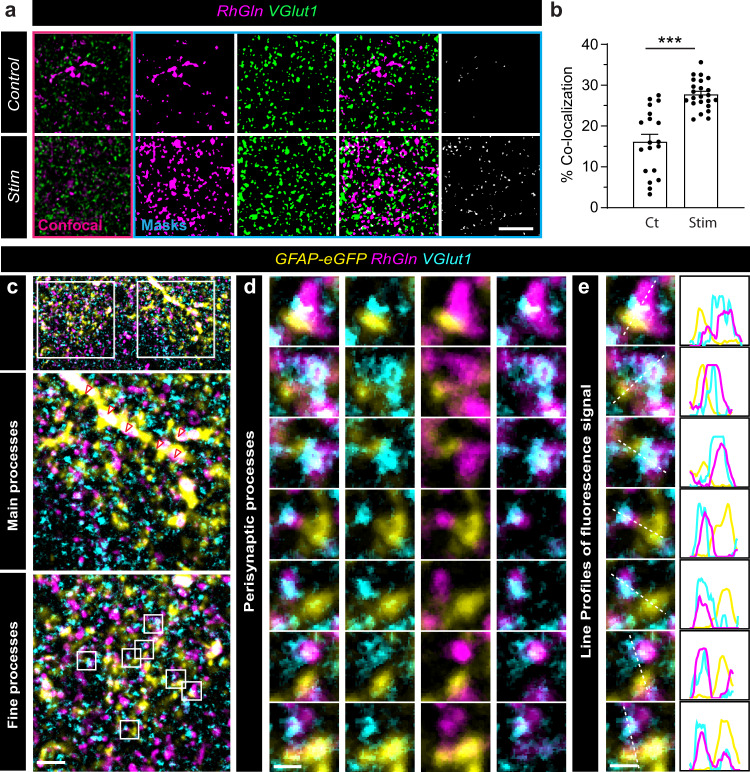
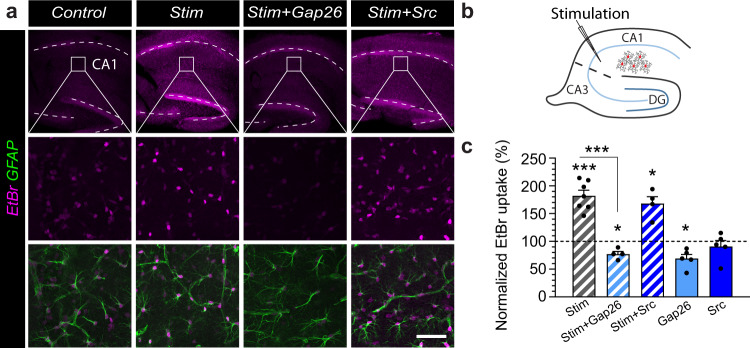
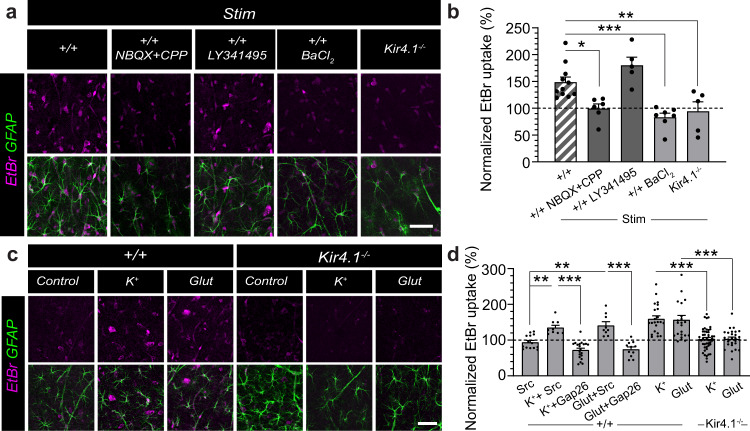
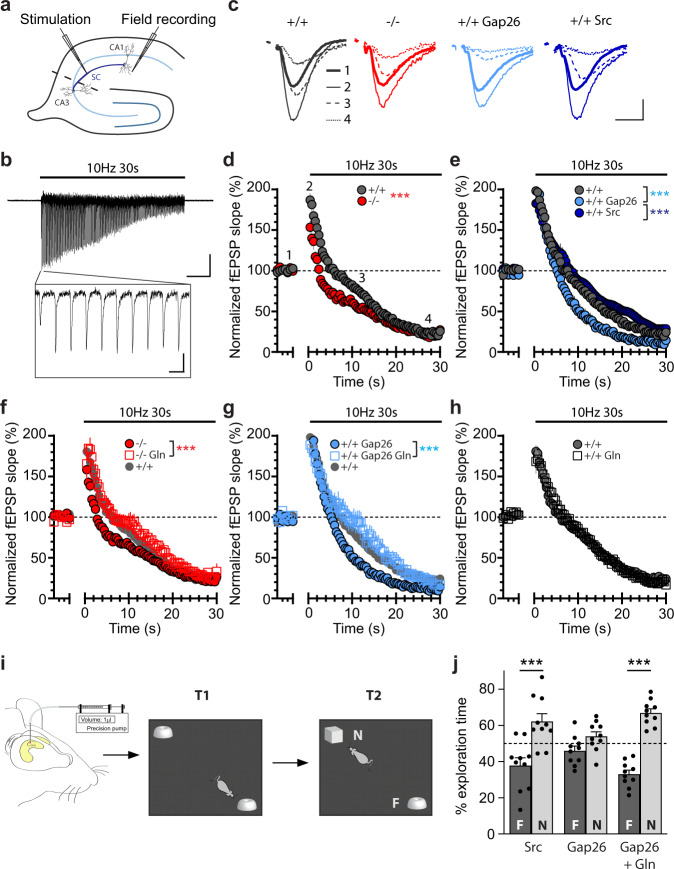
Presynaptic glutamate replenishment is fundamental to brain function. In high activity regimes, such as epileptic episodes, this process is thought to rely on the glutamate-glutamine cycle between neurons and astrocytes. However the presence of an astroglial glutamine supply, as well as its functional relevance in vivo in the healthy brain remain controversial, partly due to a lack of tools that can directly examine glutamine transfer. Here, we generated a fluorescent probe that tracks glutamine in live cells, which provides direct visual evidence of an activity-dependent glutamine supply from astroglial networks to presynaptic structures under physiological conditions. This mobilization is mediated by connexin43, an astroglial protein with both gap-junction and hemichannel functions, and is essential for synaptic transmission and object recognition memory. Our findings uncover an indispensable recruitment of astroglial glutamine in physiological synaptic activity and memory via an unconventional pathway, thus providing an astrocyte basis for cognitive processes.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Comment in
-
Role of astroglial glutamine in memory.Lab Anim (NY). 2022 Apr;51(4):99. doi: 10.1038/s41684-022-00954-9. Lab Anim (NY). 2022. PMID: 35361987 No abstract available.
References
-
- Rothstein JD, et al. Knockout of glutamate transporters reveals a major role for astroglial transport in excitotoxicity and clearance of glutamate. Neuron. 1996;16:675–686. - PubMed
-
- Danbolt NC, et al. Properties and localization of glutamate transporters. Prog. Brain Res. 1998;116:23–43. - PubMed
-
- Bak LK, Schousboe A, Waagepetersen HS. The glutamate/GABA-glutamine cycle: aspects of transport, neurotransmitter homeostasis and ammonia transfer. J. Neurochem. 2006;98:641–653. - PubMed
-
- Schousboe A, et al. Trafficking between glia and neurons of TCA cycle intermediates and related metabolites. Glia. 1997;21:99–105. - PubMed
-
- Hertz L, Dringen R, Schousboe A, Robinson SR. Astrocytes: glutamate producers for neurons. J. Neurosci. Res. 1999;57:417–428. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
