

In vivo mitochondrial base editing via adeno-associated viral delivery to mouse post-mitotic tissue
- PMID: 35136065
- PMCID: PMC8825850
- DOI: 10.1038/s41467-022-28358-w
In vivo mitochondrial base editing via adeno-associated viral delivery to mouse post-mitotic tissue
Abstract
Mitochondria host key metabolic processes vital for cellular energy provision and are central to cell fate decisions. They are subjected to unique genetic control by both nuclear DNA and their own multi-copy genome - mitochondrial DNA (mtDNA). Mutations in mtDNA often lead to clinically heterogeneous, maternally inherited diseases that display different organ-specific presentation at any stage of life. For a long time, genetic manipulation of mammalian mtDNA has posed a major challenge, impeding our ability to understand the basic mitochondrial biology and mechanisms underpinning mitochondrial disease. However, an important new tool for mtDNA mutagenesis has emerged recently, namely double-stranded DNA deaminase (DddA)-derived cytosine base editor (DdCBE). Here, we test this emerging tool for in vivo use, by delivering DdCBEs into mouse heart using adeno-associated virus (AAV) vectors and show that it can install desired mtDNA edits in adult and neonatal mice. This work provides proof-of-concept for use of DdCBEs to mutagenize mtDNA in vivo in post-mitotic tissues and provides crucial insights into potential translation to human somatic gene correction therapies to treat primary mitochondrial disease phenotypes.
© 2022. The Author(s).
Conflict of interest statement
M.M. is a co-founder, shareholder and member of the Scientific Advisory Board of Pretzel Therapeutics, Inc. P.S.-P., P.A.N., and C.D.M. provide consultancy services for Pretzel Therapeutics, Inc. The remaining authors declare no competing interests.
Figures
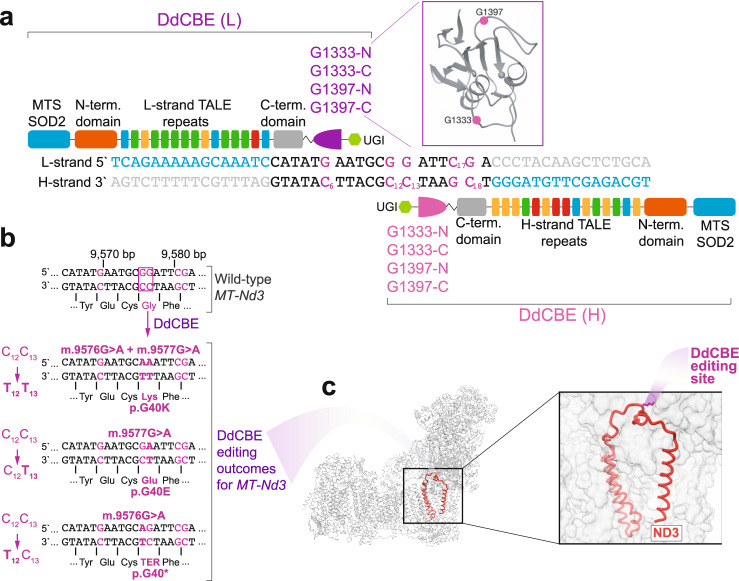
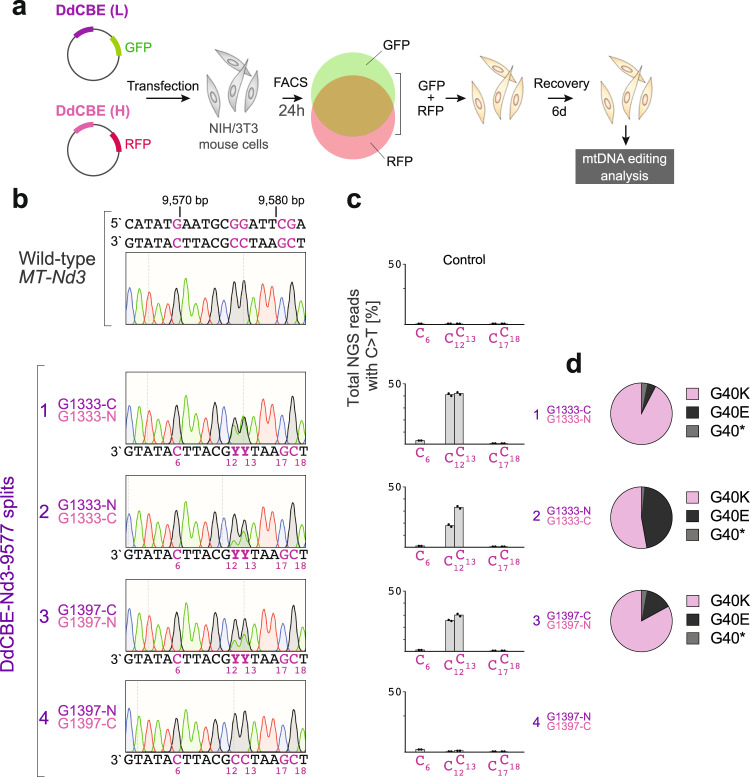
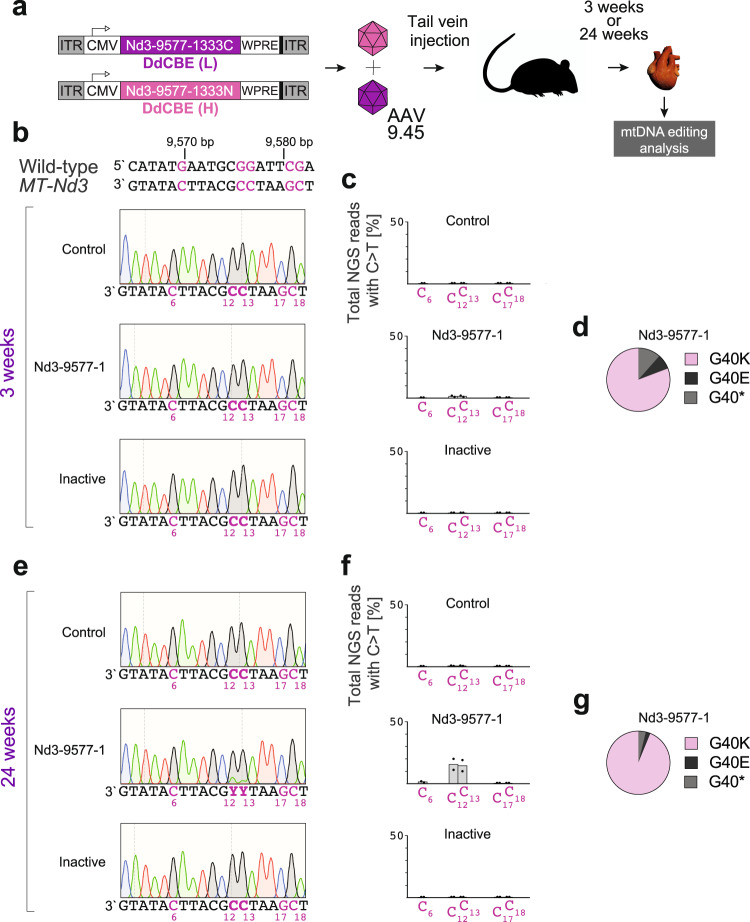
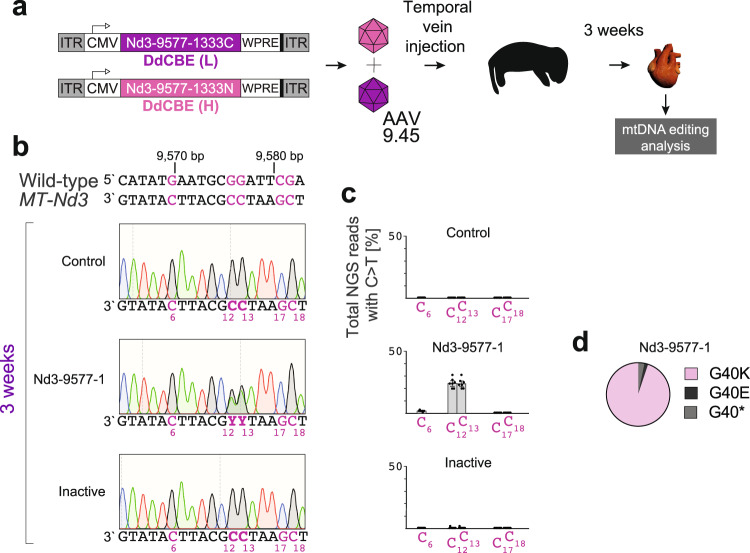
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
