

Perinatal exposure to pesticides alters synaptic plasticity signaling and induces behavioral deficits associated with neurodevelopmental disorders
- PMID: 35137321
- PMCID: PMC10547633
- DOI: 10.1007/s10565-022-09697-2
Perinatal exposure to pesticides alters synaptic plasticity signaling and induces behavioral deficits associated with neurodevelopmental disorders
Abstract
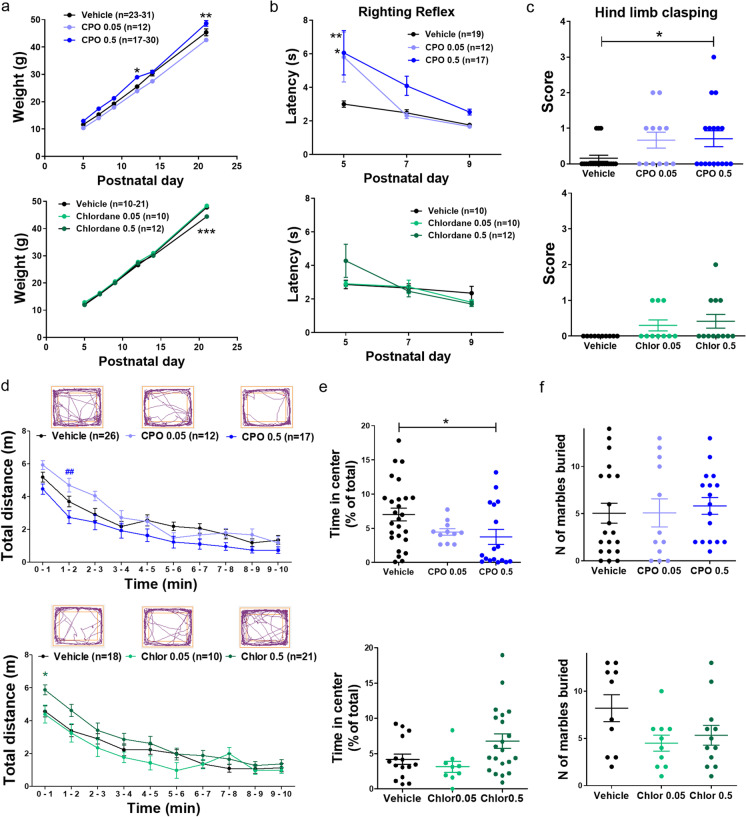
Increasing evidence from animal and epidemiological studies indicates that perinatal exposure to pesticides cause developmental neurotoxicity and may increase the risk for psychiatric disorders such as autism and intellectual disability. However, the underlying pathogenic mechanisms remain largely elusive. This work was aimed at testing the hypothesis that developmental exposure to different classes of pesticides hijacks intracellular neuronal signaling contributing to synaptic and behavioral alterations associated with neurodevelopmental disorders (NDD). Low concentrations of organochlorine (dieldrin, endosulfan, and chlordane) and organophosphate (chlorpyrifos and its oxon metabolite) pesticides were chronically dosed ex vivo (organotypic rat hippocampal slices) or in vivo (perinatal exposure in rats), and then biochemical, electrophysiological, behavioral, and proteomic studies were performed. All the pesticides tested caused prolonged activation of MAPK/ERK pathway in a concentration-dependent manner. Additionally, some of them impaired metabotropic glutamate receptor-dependent long-term depression (mGluR-LTD). In the case of the pesticide chlordane, the effect was attributed to chronic modulation of MAPK/ERK signaling. These synaptic alterations were reproduced following developmental in vivo exposure to chlordane and chlorpyrifos-oxon, and were also associated with prototypical behavioral phenotypes of NDD, including impaired motor development, increased anxiety, and social and memory deficits. Lastly, proteomic analysis revealed that these pesticides differentially regulate the expression of proteins in the hippocampus with pivotal roles in brain development and synaptic signaling, some of which are associated with NDD. Based on these results, we propose a novel mechanism of synaptic dysfunction, involving chronic overactivation of MAPK and impaired mGluR-LTD, shared by different pesticides which may have important implications for NDD.
Keywords: Contaminants; Kinase signaling; MAPK; Neurodevelopment; Psychiatric disorders; mGluR LTD.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






Similar articles
-
Translational outcomes relevant to neurodevelopmental disorders following early life exposure of rats to chlorpyrifos.J Neurodev Disord. 2020 Dec 16;12(1):40. doi: 10.1186/s11689-020-09342-1. J Neurodev Disord. 2020. PMID: 33327943 Free PMC article.
-
Chlorpyrifos-induced dysregulation of synaptic plasticity in rat hippocampal neurons.J Environ Sci Health B. 2023;58(2):100-109. doi: 10.1080/03601234.2023.2171236. Epub 2023 Feb 1. J Environ Sci Health B. 2023. PMID: 36722685
-
A Complex Interaction Between Reduced Reelin Expression and Prenatal Organophosphate Exposure Alters Neuronal Cell Morphology.ASN Neuro. 2016 Jun 30;8(3):1759091416656253. doi: 10.1177/1759091416656253. Print 2016 Jun. ASN Neuro. 2016. PMID: 27364165 Free PMC article.
-
Potential role of organochlorine pesticides in the pathogenesis of neurodevelopmental, neurodegenerative, and neurobehavioral disorders: A review.Life Sci. 2016 Jan 15;145:255-64. doi: 10.1016/j.lfs.2015.11.006. Epub 2015 Nov 5. Life Sci. 2016. PMID: 26549647 Review.
-
Pesticides and neurodevelopment of children in low and middle-income countries: A systematic review.PLoS One. 2025 Jun 11;20(6):e0324375. doi: 10.1371/journal.pone.0324375. eCollection 2025. PLoS One. 2025. PMID: 40498761 Free PMC article.
Cited by
-
The susceptibility of humans to neurodegenerative and neurodevelopmental toxicities caused by organophosphorus pesticides.Arch Toxicol. 2023 Dec;97(12):3037-3060. doi: 10.1007/s00204-023-03604-2. Epub 2023 Oct 3. Arch Toxicol. 2023. PMID: 37787774 Review.
-
The Autism Spectrum Disorder and Its Possible Origins in Pregnancy.Int J Environ Res Public Health. 2024 Feb 20;21(3):244. doi: 10.3390/ijerph21030244. Int J Environ Res Public Health. 2024. PMID: 38541246 Free PMC article. Review.
-
Chlorpyrifos residue level and ADHD among children aged 1-6 years in rural China: A cross-sectional study.Front Pediatr. 2022 Oct 14;10:952559. doi: 10.3389/fped.2022.952559. eCollection 2022. Front Pediatr. 2022. PMID: 36313880 Free PMC article.
-
Editorial: Advances in understanding the pathogenetic mechanisms of neurodevelopmental disorders and neurodegenerative disease - The environment as a putative risk factor.Front Neurol. 2023 Aug 3;14:1259772. doi: 10.3389/fneur.2023.1259772. eCollection 2023. Front Neurol. 2023. PMID: 37602235 Free PMC article. No abstract available.
-
Neurotoxic Effects of Pesticides: Implications for Neurodegenerative and Neurobehavioral Disorders.J Xenobiot. 2025 Jun 1;15(3):83. doi: 10.3390/jox15030083. J Xenobiot. 2025. PMID: 40558866 Free PMC article. Review.
References
-
- Al‐Hachim GM, Al‐Baker A. Effects of chlordane on conditioned avoidance response, brain seizure threshold and open‐field performance of prenatally‐treated mice. Br. J. Pharmacol. [Internet]. John Wiley & Sons, Ltd; 1973 Oct 1 [cited 2020 May 4];49(2):311–5. Available from: http://doi.wiley.com/10.1111/j.1476-5381.1973.tb08377.x - DOI - PMC - PubMed
-
- Albertson TE, Walby WF, Stark LG, Joy RM. The effects of lindane and long-term potentiation (LTP) on pyramidal cell excitability in the rat hippocampal slice. Neurotoxicology [Internet]. 1997 [cited 2021 Sep 13];18(2):469–78. Available from: https://pubmed.ncbi.nlm.nih.gov/9291495/ - PubMed
-
- Anway MD, Cupp AS, Uzumcu M, Skinner MK. Epigenetic Transgenerational actions of endocrine disruptors and male fertility. Science (80-. ). [Internet]. American Association for the Advancement of Science ; 2005 Jun 3 [cited 2021 Oct 25];308(5727):1466–9. Available from: https://www.science.org/doi/abs/10.1126/science.1108190 - DOI - PMC - PubMed
-
- Batista TH, Giusti-Paiva A, Vilela FC. Maternal protein malnutrition induces autism-like symptoms in rat offspring. Nutr. Neurosci. Taylor and Francis Ltd.; 2019 22(9):655–63. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
