

Chemotropism and Cell-Cell Fusion in Fungi
- PMID: 35138122
- PMCID: PMC8826960
- DOI: 10.1128/mmbr.00165-21
Chemotropism and Cell-Cell Fusion in Fungi
Abstract
Fungi exhibit an enormous variety of morphologies, including yeast colonies, hyphal mycelia, and elaborate fruiting bodies. This diversity arises through a combination of polar growth, cell division, and cell fusion. Because fungal cells are nonmotile and surrounded by a protective cell wall that is essential for cell integrity, potential fusion partners must grow toward each other until they touch and then degrade the intervening cell walls without impacting cell integrity. Here, we review recent progress on understanding how fungi overcome these challenges. Extracellular chemoattractants, including small peptide pheromones, mediate communication between potential fusion partners, promoting the local activation of core cell polarity regulators to orient polar growth and cell wall degradation. However, in crowded environments, pheromone gradients can be complex and potentially confusing, raising the question of how cells can effectively find their partners. Recent findings suggest that the cell polarity circuit exhibits searching behavior that can respond to pheromone cues through a remarkably flexible and effective strategy called exploratory polarization.
Keywords: Cdc42; GPCR; anastomosis; cell polarity; fungi; mating; pheromone; yeast.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

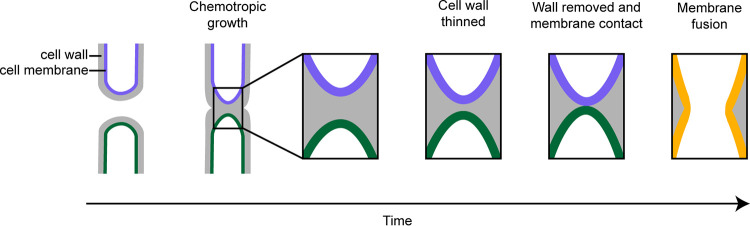

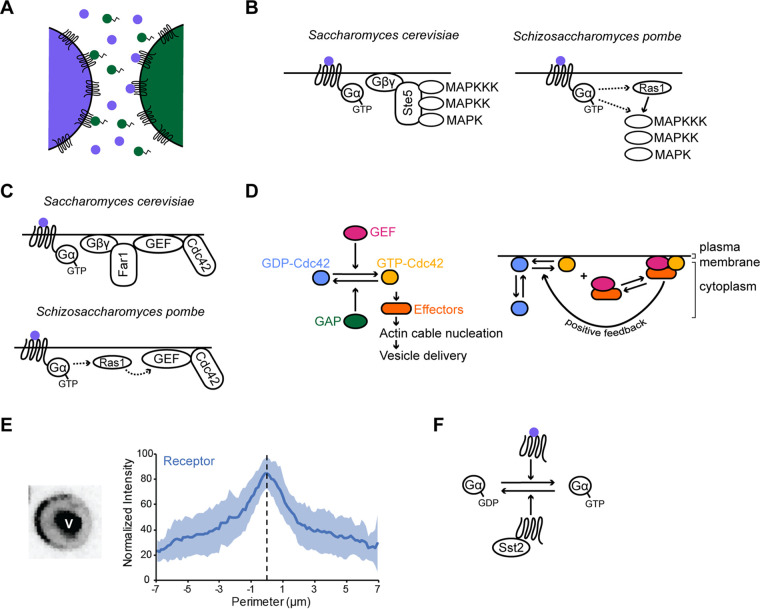



References
-
- Erental A, Dickman MB, Yarden O. 2008. Sclerotial development in Sclerotinia sclerotiorum: awakening molecular analysis of a “dormant” structure. Fungal Biol Rev 22:6–16. 10.1016/j.fbr.2007.10.001. - DOI
-
- Buller AHR. 1958. The formation of hyphal fusions in the mycelium of the higher fungi, p 27–32. In Researches on fungi: hyphal fusions and protoplasmic streaming in the higher fungi, together with an account of the production and liberation of spores in Sporobolomyces, Tilletia, and Sphaerobolus, vol 5. Hafner Publishing Co., New York, NY.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
